Sinh học cơ thể thực vật
CHƯƠNG
3
SỰ
PHÁT TRIỂN VÀ CÁC QUÁ TRÌNH
ÐIỀU
HÒA SINH TRƯỞNG Ở THỰC VẬT
Trong chu kỳ đời sống ở thực
vật hột kín, kết thúc một chu kỳ là
sự hình thành hột, trong đó có chứa một
thế hệ bào tử thực vật mới.
Bắt đầu cho một chu kỳ đời
sống mới là khi hột nẩy mầm và hình thành
cây con.
Nhiều
hột phải trải qua một thời gian miên
trạng dài trước khi nẩy mầm.
Các yếu tố của môi trường như nước,
nhiệt độ và oxy... cũng ảnh hưởng trên
sự nẩy mầm của hột.
|
|
a.
Sự nẩy mầm của hột
Sự
nẩy mầm của hột bắt đầu khi
hột hấp thu rất nhiều nước và tăng
thể tích lên một cách đáng kể, có khi đến
200%. Kết quả
của sự hấp thu nước này làm cho phôi
giải phóng gibberellin, và đây là yếu tố
cảm ứng để tổng hợp một số các
enzim thủy giải trong đó có cả amylaz.
Những enzim này thủy phân những chất
dự trử trong phôi nhũ, cung cấp năng lượng
cho sự tăng trưởng của phôi.
Sự biến dưỡng của phôi tăng,
sự gia tăng này làm cho tế bào bắt đầu
phân cắt, tổng hợp thêm tế bào chất
mới, và tế bào gia tăng kích thước nhờ
sự hấp thu nước.
Phôi tăng trưởng làm bung vỏ hột ra và
nhanh chóng hình thành một cây con, có rễ và thân phân
biệt.
 |
Khi hột nẩy mầm, trục hạ diệp được mọc ra trước tiên. Trục hạ diệp mọc xuống theo chiều trọng lực, dù hột nằm theo hướng nào. Cùng lúc đó trục thượng diệp bắt đầu phát triển nhanh chóng, rễ mầm ở phần cuối của trục hạ diệp, tạo ra một hệ thống rễ con để gắn vào trong đất và hấp thu nước và muối khoáng. Ở một số cây song tử diệp, phần trên của trục hạ diệp mọc dài ra thành dạng hình vòm, mọc ngược lên và chui ra khỏi mặt đất (Hình 1A). Khi trục hạ diệp lộ ra ngoài không khí, nó mọc thẳng lên, tử diệp và trục thượng diệp được đưa ra khỏi mặt đất. Sau đó trục thượng diệp bắt đầu mọc dài ra. Ðây là kiểu nẩy mầm thượng địa.
Những cây Song tử diệp khác, thí dụ như
đậu Hà lan, Nhản có một kiểu nẩy
mầm hơi khác, ở những cây này, trục hạ
diệp không mọc thành hình vòm và tử diệp không
được đưa lên khỏi mặt đất.
Thay vào đó là trục thượng diệp
bắt đầu mọc dài ra ngay sau khi hệ thống
rễ con bắt đầu được hình thành; nó
luôn luôn mọc thẳng đứng va ìchẳng bao lâu
nhô ra khỏi mặt đất.
Kiểu nẩy mầm này tương tự như
ở hột Lúa, Bắp... thuộc các cây đơn
tử diệp, chỉ có một tử diệp, nhưng
giàu phôi nhũ (Hình 1B). Ðây
là kiểu nẩy mầm hạ địa.
Ở Bắp trục thượng diệp bắt
đầu dài ra ngay sau khi hệ thống rễ
được thành lập.
Thân non được diệp tiêu (lá đầu
tiên hình ống) bao bọc.
b. Sự
phát triển của cây con
Ðầu
tiên cây con tăng trưởng hơi chậm, nhưng
sau đó tăng trưởng với một tốc
độ nhanh hơn trong một thời gian dài hơn và
cuối cùng chậm lại và có thể dừng tăng
trưởng khi cây sắp trưởng thành.
Ở những cây đa niên, sự tăng trưởng
tiếp tục xảy ra trong suốt đời sống
của cây, trong khi ở những cây nhất niên như
các cây Ðậu, cây
Củ cải... tăng trưởng ngừng lại khi
cây trưởng thành và cây chết đi sau một mùa
sinh trưởng. Sự
tăng trưởng của rễ và thân của cây con
có được là nhờ sự phân cắt và sự
tăng dài của tế bào.
Hai hoạt động này chịu ảnh hưởng
của nhiều hormon sinh trưởng khác nhau, đặc
biệt là auxin, gibberellin và cytokinin.
Ở những cây chỉ có mô sơ cấp thì
sự phân cắt tế bào và sự tăng dài của
tế bào tùy thuộc vào sự hoạt động
của hai mô phân sinh ngọn rễ và ngọn thân.
|
|
|
|

a.
Sự tăng trưởng của rễ
Sự hoạt động của mô phân sinh ngọn rễ làm cho rễ tăng trưởng. Mô phân sinh rễ được bảo vệ bởi một chóp rễ hình nón, gồm một khối tế bào không phân cắt được. Khi rễ mọc dài ra và đầu rễ mọc sâu vào trong đất thì một số tế bào ở mặt ngoài của chóp rễ có thể bị tổn thương và sau đó được thay thế bằng những tế bào mới do sự phân cắt tế bào của mô phân sinh ngọn. Ngay sau của chóp rễ là vùng mô phân sinh ngọn rễ, vùng này ngắn và gồm những tế bào nhỏ có khả năng phân chia tích cực. Phần lớn các tế bào mới được tạo ra nằm xa chóp rễ. Mô phân sinh tiếp tục phân cắt cho tế bào mới và đầu rễ tiếp tục mọc sâu vào trong đất. Chính các tế bào được tạo ra từ mô phân sinh này sẽ thành lập mô sơ cấp cho rễ (Hình 2).
Khi
các tế bào được mới được
đẩy ra khỏi vùng mô phân sinh ngọn, do số lượng
tế bào tăng lên sự phân cắt chậm lại
thì sự gia tăng kích thước tế bào là quá trình
chính. Phần lớn
sự tăng kích thước làm rễ tăng trưởng
chiều dài nhiều hơn là chiều rộng.
Sự tăng dài của tế bào chịu tác
động của các hormon mà đặc biệt là
auxin và gibberellin. Vùng
tế bào tăng dài nhiều nhất là vùng ngay sau vùng
mô phân sinh và thường dài chỉ vài milimet.
Kế tiếp là vùng tế bào trưởng thành,
nơi đây tế bào bắt đầu trưởng
thành và có hình thành dạng đặc trưng.
Vùng này dễ nhận biết nhờ các lông hút
được mọc dài ra từ những tế bào
biểu bì.
b.
Sự tăng trưởng của thân
|
|

|
|
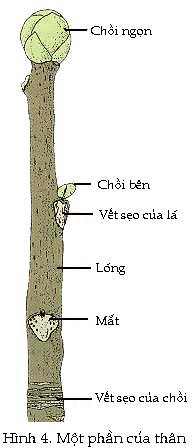
Ở đỉnh của thân là một chuỗi
những lóng chưa được mọc dài ra.
Những khối sơ khởi của lá rất
nhỏ ngăn cách các lóng uốn cong; các khối sơ
khởi già hơn, to hơn của lá bao lấy các
khối sơ khởi trẻ hơn, nhỏ hơn ở
bên trong. Cấu trúc
gồm mô phân sinh ngọn và các lóng chưa được
tăng dài được bao bọc trong các khối sơ
khởi của lá được gọi là chồi
(bud). Ở những cây
tăng trưởng theo mùa thì chồi được
bảo vệ bởi những vảy, là những lá
biến đổi mọc từ dưới đáy
của chồi. Vào mùa
xuân, khi các chồi ngũ này nở ra, thì các vảy
che chở rụng đi và những lóng chứa bên trong
các chồi bắt đầu tăng dài một cách
nhanh chóng. Do đó các
lóng sẽ dần dần được tách xa nhau ra,
sự phân cắt tế bào xảy ra ở khối sơ
khởi của lá và tạo ra lá non.
Trước khi lá được hình thành một
cách hoàn chỉnh, một u nhỏ của mô phân sinh thường
mọc ra ở giữa đáy lá và lóng.
Mỗi một vùng mô phân sinh mới này sẽ
tạo ra một chồi bên (lateral, axillary bud) có đặc
điểm tương tự như chồi ngọn (Hình
4). Sự tăng dài
của các lóng của chồi bên trong mùa sinh trưởng
kế tiếp sẽ tạo ra nhánh.
*
Sự chuyên hóa của tế bào
Tất
cả những tế bào
mới được sinh ra từ mô phân sinh thì cơ
bản giống nhau, chúng
sẽ trở thành các loại mô khác nhau.
Quá trình tế bào thay đổi từ những hình
dạng chưa trưởng thành đến trưởng
thành gọi là sự chuyên hóa (differentiation).
Trong
sự tăng trưởng của rễ và thân, tế
bào bắt đầu chuyên hóa thành các loại mô khác
nhau khi chúng vẫn còn ở trong vùng mô phân sinh (Hình 2).
Sau khi sự phân cắt tế bào và sự tăng
dài của tế bào đã hoàn tất, tế bào
bắt đầu trưởng thành có hình dạng
nhất định. Ở
lát cắt ngang có thể phân biệt được ba
vùng đồng tâm ngay sau mô phân sinh của rễ đó
là lớp tiền bì (protoderm), kế tiếp là một
vùng mô căn bản dày nằm ngay dưới tiền
bì và trong cùng là mô tiền dẫn truyền (provascular
tissue) gồm những tế bào dài (Hình 5A).
Ngay trong phôi, tiền bì ngoài trở thành biểu bì,
mô căn bản trở thành vỏ và nội bì,
phần trong cùng tạo ra mô dẫn truyền sơ
cấp, chu luân và tượng tầng libe gỗ (Hình 5B
và C). Sự chuyên hóa
trong thân đang tăng trưởng cũng theo cách tương
tự ngoại trừ có hai vùng mô căn bản,
một vùng nằm giữa tiền bì và trụ tiền
dẫn truyền sẽ tạo ra vỏ và nội bì, và
một vùng thứ hai nằm trong trụ tiền dẫn
truyền sẽ trở thành lỏi.
Sự tăng trưởng theo đường kính
của rễ và thân tùy thuộc vào sự thành lập
mô thứ cấp do sự hoạt động của
những mô phân sinh bên, đặc biệt là tượng
tầng libe gỗ. Dưới
ảnh hưởng của auxin, những tế bào
mới được tạo ra ở phía ngoài của tượng
tầng sẽ chuyên hóa thành mô libe thứ cấp, trong
khi đó những tế bào mới được
tạo ra ở phía trong của tượng tầng
sẽ tạo nên mô gỗ thứ cấp (Hình 5D và E).

|
|
|
|

Thực vật có xu hướng mọc về hướng có ánh sáng. Ðặt một chậu cây trong phòng, cây sẽ mọc cong hướng về phía cửa sổ, nếu xoay cây hướng vào trong, sau một thời gian ngắn cây lại mọc hướng về phía cửa sổ (Hình 6). Hiện tượng cây đáp ứng lại với ánh sáng bởi sự xoay này được gọi là quang hướng động (phototropism) của thực vật. Thực vật còn có các tính hướng động khác như địa hướng động (gravitropism) là đáp ứng của cây hướng theo chiều của trọng lực, thủy hướng động (hydrotropism) đáp ứng với nước.
Ðáp ứng này là do sự sinh trưởng chuyên hóa; một phía của thân cây hay rễ mọc nhanh hơn phía bên kia, làm cho cây cong đi. Thân có quang hướng động dương, xoay về hướng có ánh sáng; rễ thì ngược lại, có quang hướng động âm, xoay tránh ánh sáng. Ý nghĩa thích nghi của quang hướng động ở thân là xoay thân để lá nhận được ánh sáng tối đa cần thiết cho sự quang hợp.

Một
trong các thí nghiệm đầu tiên tìm hiểu quang hướng
động của cây của
Charles Darwin với con trai là Francis khoảng 1880.
Ðối tượng thí nghiệm của ông là diệp
tiêu (coleoptile) của cỏ, diệp tiêu tăng trưởng
chủ yếu do tế bào tăng dài và có tính quang
hướng động thuận (+) (Hình 7A). Ông cho
thấy rằng nếu ngọn diệp tiêu được
đậy lại bằng một chụp đen thì
ngọn không mọc cong hướng về nơi có ánh
sáng (Hình 7B và C), trong khi những cây kiểm chứng
với ngọn diệp tiêu trần hay bao lại
bằng một cái chụp trong suốt cho ánh sáng đi
qua thì diệp tiêu sẽ mọc cong hướng về
nơi có ánh sáng (Hình 7D). Họ
cũng nhận thấy thân của diệp tiêu
được bọc lại bằng một ống
đen chừa ra phần ngọn, thì diệp tiêu cũng
mọc cong (Hình 7E). Như
vậy có lẻ ngọn diệp tiêu có vai trò then
chốt trong sự quang hướng động của cây.
Ðiều này được khẳng định
bởi thí nghiệm diệp tiêu bị cắt ngọn
(B) thì không thể mọc cong được.
Diệp tiêu đối chứng (A) với ngọn
nguyên vẹn mọc cong bình thường. Darwin kết
luận rằng ánh sáng có tác động trên phần
đỉnh ngọn của diệp tiêu và có chất gì
đó được vận chuyển từ trên
xuống làm phần dưới ngọn diệp tiêu
mọc cong đi.
II.
CÁC HORMON TĂNG TRƯỞNG CỦA THỰC VẬT
Thực vật phản ứng lại với
những kích thích bên ngoài như ánh sáng, nhiệt độ,
trọng lực, độ dài của ngày... nhưng vì
chúng không có hệ thần kinh và không di chuyển như
động vật do đó những phản ứng
của chúng chậm và không rõ ràng.
Thực vật thường phản ứng lại
với những kích thích bằng sự tăng trưởng.
Thí dụ, thân và lá mọc hướng về ánh
sáng, rễ mọc hướng theo chiều của
trọng lực, thân mọc ngược chiều
với chiều của trọng lực.
Nhiều phản ứng có thể xảy ra nhanh, thí
dụ sau một vài phút cây bị mất nước, cây
bắt đầu héo, khí khẩu đóng lại, và
một số gen bắt đầu hoạt động,
phiên mã, giải mã nhanh chóng để tạo ra protein
giúp cho cây chống chịu ảnh hưởng của
khô hạn.
Cây trao đổi thông tin giữa các tế bào
chủ yếu là nhờ các hóa chất.
Khi cây nhận một kích thích, hormon, một hóa
chất kiểm soát đặc biệt được
tạo ra. Hormon có
thể hoạt động tại chỗ, hay có thể
được vận chuyển đến một
tế bào điểm ở một phần khác của cây,
ảnh hưởng quá trình sinh lý nơi đó.
Hormon hoạt động như người
đưa tin giữa các tế bào và có hiệu qủa
với một nồng độ rất thấp.
Mỗi hormon chỉ ảnh hưởng đến
một quá trình sinh lý chuyên biệt.
Hiện nay đã biết 5 nhóm hormon: auxin, gibberellin, cytokinin, acid abscisic và ethylen.
| 1. Auxin |
|
a.
Vai trò của auxin trong sự tăng dài của tế bào
|
|

Auxin
có vai trò quan trọng trong sự kiểm soát sự tăng
dài của tế bào. Vì
tế bào thực vật có vách bao bọc, nên tế bào
chỉ có thể tăng trưởng được khi
vách có thể được kéo dài ra.
Vách được cấu tạo bởi phần
lớn là đường đa mà thành phần chính là
celluloz. Ở vách sơ
cấp, celluloz hiện diện dưới dạng
những sợi dài liên kết với các đường
đa khác để tạo ra một mạng lưới
(Hình 8). Khi tăng trưởng
các liên kết có thể bị đứt tạm
thời, do đó vách tế bào trở
nên đàn hồi (nới rộng) hơn và
những vật liệu mới được chen vào.
Auxin có vai trò chính trong cả hai quá trình trên.
Aính
hưởng của auxin trên sự đàn hồi hóa
của vách xảy ra nhanh chóng (khoảng 10 phút).
Theo giả thuyết về sự tăng trưởng
acid, auxin có ảnh hưởng gián tiếp bằng cách
acid hóa vách tế bào. Auxin
điều tiết pH của vách tế bào bằng
cảm ứng sự vận chuyển tích cực ion H+
từ tế bào chất vào vách tế bào.
pH acid hoạt hóa enzim trong vách, bẻ gảy các liên
kết chéo giữa các sợi celluloz.
Các sợi lỏng lẻo làm tế bào trở nên
đàn hồi hơn.
|
|

Môi
trường trong tế bào ưu trương hơn so
với dịch lỏng bên ngoài tế bào, và nước
đi vào trong tế bào bởi sự thẩm thấu,
tạo ra một áp suất trương bên trong tế bào.
Khi vách trở nên đàn hồi hơn chúng dễ
dàng được kéo căng ra.
Nước tiếp tục đi vào tế bào
bằng sự thẩm thấu, làm gia tăng thể tích
tế bào (Hình 9), nhưng sự gia tăng kích thước
này không có sự tổng hợp ra tế bào chất
mới.
Aính hưởng của auxin trên sự căng vách,
thêm vào các vật liệu mới vào thì chậm hơn
vì vách này tùy thuộc vào sự kích động
của một số gen, sự tổng hợp những
mARN chuyên biệt, để tổng hợp các enzim xúc
tác tạo thêm những đơn vị đường
đa vào vách tế bào. Khi
tế bào được cung cấp auxin, nó hoạt hóa
bơm ion H+ trên màng sinh chất (Hình 9A).
Ion H+ được vận chuyển tích cực
từ tế bào chất vào trong vách.
Sự gia tăng ion H+ hoạt hóa enzim bẻ
gảy một số liên kết chéo giữa các
đường đa cấu tạo vách và vách trở
nên mềm dẻo hơn (Hình 9B).
Vì nước vào không bào càng lúc càng nhiều vách
sẽ bị căng ra, nhưng chỉ theo một hướng.
Hầu như không có sự tổng hợp tế
bào chất mới trong suốt thời gian tăng trưởng
theo kiểu này. Sự
gia tăng thể tích tế bào là do sự phát
triển của không bào (Hình 9C).
Các không bào nhỏ hòa vào nhau thành một không bào
to duy nhất; ở tế bào trưởng
thành dãy tế bào chất ở ngoại biên
chỉ chiếm 10% thể tích tế bào (Hình 9D).
b.
Vai trò trung gian của auxin trong quang hướng động
của cây

Năm
1926 Frits Went (người Hà lan), làm thí nghiệm cắt
ngọn của diệp tiêu và đặt ngọn này lên
một khối agar trong chừng một giờ (Hình 10) và
được chiếu sáng.
Sau đó đặt khối agar không có diệp tiêu
này lên phần diệp tiêu còn lại.
Phần diệp tiêu này tiếp tục tăng trưởng
mọc cong về phía chiếu sáng và nếu đặt
khối agar lệch trục diệp tiêu thì diệp tiêu
cũng mọc cong ngay cả trong tối.
Khối agar kiểm chứng không gây một
hiệu qủa nào. Rõ
ràng rằng có một chất kích thích sinh trưởng
đã khuếch tán từ phần ngọn diệp tiêu vào
khối agar và khi đặt khối agar này lên phần
diệp tiêu còn lại thì chất này được di
chuyển xuống và kích thích sự tăng dài.
Went gọi chất này là auxin (tiếng Hy lạp có
nghĩa là tăng trưởng).

Acid indoleacetic (AIA)
Tryptophan
(Trp)
Hình
11. AIA và Tryptophan
Có nhiều hóa chất, một số được
tìm thấy trong cây, một số tổng hợp trong phòng
thí nghiệm, có tác dụng như thí nghiệm của
Went và được gọi chung là auxin.
Một chất được nghiên cứu
nhiều nhất là acid indolacetic (IAA), đã được
phân lập từ nhiều nguồn thiên nhiên.
Acid indolacetic là chất hóa học tương
tự tryptophan và được tổng hợp từ nó
(Hình 11).
Các thí nghiệm trên cho thấy ngọn diệp tiêu
phóng thích auxin, chất này di chuyển xuống và kích
thích sự tăng dài của tế bào trong diệp tiêu.
Tuy nhiên, cơ chế phản ứng của quang hướng
động vẫn chưa biết rõ.
Biết rằng khi ánh sáng tác động từ
một phía ngọn cây, nó làm giảm sự cung cấp
auxin cho cây ở phía đó và tăng ở phía ở
trong tối. Ðồng
thời chứng tỏ rằng có sự vận
chuyển tích cực của auxin trong ngọn từ
phần được chiếu sáng sang phần tối.
Kết quả là bên phía được chiếu sáng
cây tăng trưởng chậm hơn phía bên tối, và
sự tăng trưởng bất xứng này làm cho cây
mọc cong về phía sáng.
Các nhà sinh lý thực vật vẫn chưa
khẳng định được bằng cách nào
ngọn cảm nhận được ánh sáng hay có các
thụ thể nào tham gia. Rõ
ràng là ánh sáng gây ra sự phân phối lại auxin
ở ngọn, tích tụ ở phía thân không có ánh sáng.
Không giống hầu hết những hormon thực
vật, auxin không được vận chuyển theo mô
dẫn truyền mà đi xuống từ tế bào này
sang tế bào khác, sự di chuyển một chiều và
có lẻ do sự vận chuyển tích cực và đòi
hỏi năng lượng. Ở
thân, auxin di chuyển từ đỉnh ngọn thân
xuống phần gốc, trong khi ở rễ, sự di
chuyển hướng từ ngọn rễ đến thân.
c.
Vai trò trung gian của auxin trong địa hướng
động của cây
Nếu
đặt một chậu cây nằm ngang trong vài
giờ, sau đó thấy thân bắt đầu mọc
cong lên (Hình 12). Ðó là
địa hướng động âm (nghịch) thân
mọc ngược lại hướng của trọng
lực. Trong khi đó
thì rễ mọc hướng xuống theo chiều
trọng lực. Thực
vật cũng như nhiều sinh vật đa bào, có
những tế bào có thể cảm nhận được
sự thay đổi vị trí và đáp ứng với
trọng lực. Ở
thân, những thể tiếp nhận trọng lực này
ở trong mô phân sinh ngọn của thân, và ở
rễ, chúng là những tế bào nằm ở phần
trung tâm của chóp rễ.
|
|

Herman
Dolk (người Hà lan) khám phá ra rằng nồng độ
auxin ở phía dưới của thân nằm ngang tăng,
trong khi ở phía trên lại giảm.
Sự phân bố không đều của auxin này kích
thích tế bào ở phía dưới tăng trưởng
nhanh hơn những tế bào ở phía trên và vì
thế thân mọc cong lên. Rễ
thì ngược lại, mọc hướng theo chiều
trọng lực. Khi
đặt rễ nằm ngang phần các tế bào
ở phía trên mọc nhanh hơn những tế bào
ở phía dưới, do đó rễ mọc hướng
xuống (Hình 13).
|
|

Những
tế bào ở vùng giữa của chóp rễ được
cho rằng cảm nhận được kích thích
của trọng lực. Các
tế bào này có chứa nhiều bột lạp.
Bột lạp và những bào quan khác đáp
ứng với trọng lực nên xếp ở đáy
của tế bào, và trong một vài giờ, rễ
bắt đầu mọc cong xuống.
Nhiều nhà thực vật tin rằng bột
lạp hoạt động như một thể cảm
nhận trọng lực, nhưng những người khác
như Randy Wayne (ở Ðại học Cornel) thì không tin
như thế. Theo Wayne
thì phải có những thụ thể đặc
biệt nằm ở đáy của những tế bào
cảm nhận được trọng lượng toàn
tế bào làm co mặt dưới tế bào và dãn
mặt trên. Tất nhiên
tất cả những cơ chế trên còn chưa
hiểu rõ.
d.
Auxin ngăn cản sự tăng trưởng của
chồi bên
Auxin
cũng có ảnh hưởng ngăn cản sự tăng
trưởng của chồi bên ở nhiều cây làm
cho chúng mọc chậm hay không mọc được.
Auxin được tạo ra từ chồi
ngọn đi xuống thân, ngăn cản sự phát
triển của chồi bên, cùng lúc kích thích sự tăng
dài của thân chính. Như
thế chồi ngọn có ưu thế ngọn (apical
dominance) so với các chồi bên và dồn năng lượng
để thân chính mọc cao so với các nhánh .
Các cành ở xa ngọn có thể tăng trưởng
được vì hiêu quả ngăn cản của auxin
không còn. Nếu
chồi ngọn được cắt bỏ đi,
một số chồi bên phía trên sẽ tăng trưởng
tạo ra nhánh và khi đó chồi ngọn của nhánh
lại ức chế các chồi bên dưới nhánhï.
Người trồng cây cảnh và hoa thường
cắt bỏ chồi ngọn và lá non để cây có
nhiều nhánh và nhiều hoa hơn.
e.
Các ứng dụng trong thương mại của auxin
Auxin
ngăn cản trái và lá rụng, do đó, nếu phun
auxin lên trái sẽ giữ được trái trên cây
đến lúc muốn thu hoạch.
Phun các chất chống auxin (auxin antagonist), thường
có tác dụng ngược lại.
Thí dụ, trước khi thu hoạch bông vải
người ta phun các anti-auxin
này để cảm ứng sự thành lập lớp
tế bào gây rụng đáy lá, làm lá rụng sớm,
nhờ thế thu hoạch trái bằng cơ giới
sẽ dễ dàng hơn.
2,4-D
là một hóa chất gần giống auxin, được
tổng hợp và được sử dụng rộng
rải
để diệt cỏ có lá rộng trong các đồng
cỏ. Cây song tử
diệp thường nhạy cảm với chất này
hơn những cây đơn tử diệp, do đó
với một nồng độ thấp của 2,4-D có
thể diệt cỏ dại song tử diệp mà không
ảnh hưởng đến cây đơn tử
diệp.
Chất
độc màu da cam, cũng là một auxin tổng
hợp được quân đội Mỹ khai quang
rừng trong chiến tranh ở Việt Nam gây ảnh hưởng
lâu dài lên sức khỏe con người và đặc
biệt có lẻ là nguyên nhân làm gia tăng của
bệnh ung thư.
| 2. Gibberellin |
|
Người Nhật từ lâu rất quen thuộc với một thứ bệnh trên cây Lúa, cây cao một cách bất thường, nhưng lại không sống được đến khi cây trưởng thành. Năm 1926 E.Kurosawa (người Nhật) tìm thấy những cây này có nhiễm một loài nấm mốc tên Gibberella fugikuroi. Ông chứng minh rằng khi nấm nhiễm vào cây con, chúng cũng tăng trưởng vơi những triệu chứng tương tự như trên. Ông cũng nhận thấy những triệu chứng tương tự khi cho cây nhiễm chất trích từ nấm mốc này và ngay cả chất trích từ môi trường đã nuôi cấy nấm mốc này trên đó. Năm 1930 người ta đã phân lập và kết tinh một chất từ Gibberella nay được gọi là gibberellin. Trong vòng 30 năm trở lại đây có hơn 70 chất khác nhau được phân lập từ nấm mốc và nhiều thực vật có hoa cũng được xếp vào gibberellin (GAs). Gibberellin thường được sử dụng trong các thí nghiệm là GA3 hay acid gibberellic, những dạng hoạt động có lẻ là GA1.
a.
Tác động của gibberellin
|
|

GA1
gây ra sự tăng dài nhanh chóng của thân bằng cách
kích thích sự phân cắt tế bào và sự tăng dài
của tế bào (Hình 14). Cơ
chế tác động chưa được hiểu rõ,
nhưng tác động khác auxin ở chỗ là không có
sự tham dự của chất có tính acid gây sự tăng
trưởng.
Dù auxin và gibberellin đều kích thích sự tăng
dài của thân nhưng ảnh hưởng của chúng
trên cây khá khác biệt. Vì
gibberellin được vận chuyển qua mô gỗ và
mô libe, trong khi auxin được di chuyển từ
tế bào này sang tế bào khác một chiều.
Gibberellin có ảnh hưởng trên toàn bộ
của cây chứ không phải trên từng vùng như
ở auxin.
Gibberellin
được tạo ra trong những mô còn non của
thân và trong hột đang phát triển.
Có nhiều loại gibberellin nhưng tất cả
đều có cùng một lộ trình sinh tổng hợp
như những hormon steroid của động vật có
xương sống như estrogen và testosterol, và cũng
giống như chúng gibberellin hòa tan trong lipid.
Do đó chúng dễ dàng đi xuyên qua màng tế bào
và có lẻ làm cho gen đặc biệt nào đó
hoạt động hay ngưng hoạt động.
Bên cạnh kích thích sự phân cắt và tăng dài
của tế bào, chúng có thể cảm ứng sự
nẩy mầm ở một số hột, cảm
ứng phôi ở một số hột sinh ra amylaz để
thủy phân tinh bột dự trử, cảm ứng
nhiều cây tăng trưởng nhanh chóng và trổ hoa
và kích thích sự phát triển của trái ở
một số loài.
b.
Ứng dụng thương mại của gibberellin
Một
trong những ứng dụng quan trọng là dùng để
tăng sản lượng mạch nha từ lúa mạch
dùng làm rượu bia. GAs
được sử dụng để làm nẩy
mầm hột lúa mạch gia tăng tạo ra enzim
thủy giải những chất dự trử trong
hột thành acid amin và đường để thành
mạch nha. Có nhiều ứng dụng thực tiển
của gibberellin trong trồng trọt.
GAs cũng được sử dụng để
làm tăng kích thước và chất lượng
của nho không hột và cũng được dùng
để kích thích sự tăng trưởng của
một số trái. Một
ứng dụng khác làm cho Cam Chanh chậm chín.
Sử dụng GAs trong tháng 11 và 12 nhằm kéo dài
ngày thu hoạch, các tháng hè là lúc mà nhu cầu rất
cao trong khi lượng cung cấp thường là không
đủ.
Ngoài
gibberellin, cytokinin cũng có vai trò chính trong sự phân
cắt tế bào. Cytokinin
được tạo ra trong ngọn rễ và trong
hột đang phát triển và cytokinin được
vận chuyển qua mô gỗ từ rễ lên thân.
Tác động của cytokinin lên sự tăng trưởng
của tế bào trong môi trường nuôi cấy mô
lệ thuộc vào sự hiện diện đồng
thời của auxin. Thật
vậy, tỉ lệ giữa cytokinin và auxin có ý nghĩa
quan trọng trong việc xác định sự chuyên hóa
của tế bào mới. Khi
trong môi trường nuôi cấy cytokinin nhiều hơn
auxin thì thân và lá được tạo ra trong khi auxin
nhiều hơn thì sự tăng trưởng của
rễ bắt đầu. Sự
cân đối giữa hai loại hormon này xác định
loại mô sẽ phát triển; do đó chúng kiểm soát
hình thái của cây.
Trong
một cây phát triển bình thường, cytokinin và
auxin có hoạt động phối hợp trong một
số trường hợp, nhưng trong một số trường
hợp khác lại có tác động đối
nghịch nhau. Thí
dụ, chúng có tác động hổ trợ nhau trong
sự thúc đẩy tế bào phân cắt nhưng
lại có tác động đối nghịch về
ảnh hưởng sự tăng trưởng của
chồi bên. Cả hai
hormon đều ảnh hưởng lên sự tăng trưởng
của tế bào nhưng auxin chỉ kích thích sự tăng
dài trong khi cytokinin thúc đẩy sự phân cắt
tế bào.
Ngoài ra, cytokinin còn kích thích sự biến đổi
của những lạp còn non thành lục lạp; giúp
chồi bên tránh bớt được sự ức
chế của chồi ngọn, làm chậm sự lão hóa,
đặc biệt là đối với lá.
Hormon
acid abscisic đầu tiên được xem là kiểm
soát sự rụng lá ở nhiều cây như tên
của nó (abscission: tách ra). Nó
cũng được xem là chất cảm ứng miên
trạng của chồi và hột vào mùa thu để
đợi đến mùa xuân.
Ngày nay, người ta biết vai trò kiểm soát
sự rụng lá là không quan trọng, mà dường như
có vai trò trong miên trạng của hột, ít nhất là
ở một số loài.
Mặc
dù từ lâu ta acid abscisic được xem là yếu
tố ngăn cản sự tăng trưởng, nhưng
trong nhiều trường hợp nó thật sự
cảm ứng tăng trưởng và phát triển.
Thí dụ chúng thúc đẩy sự vận
chuyển những sản phẩm quang hợp cho phôi phát
triển trong hột và cảm ứng sự tổng
hợp protein để dự trử trong hột.
Ngoài ra, chúng còn giúp các khí khẩu đóng
lại khi trời khô. Chỉ
vài phút sau khi cây bắt đầu héo, nồng độ
acid abscisic trong lá tăng lên 10 lần.
Hormon này thật sự gây ra một số đáp
ứng, bao gồm sự đóng các khí khẩu giúp cây
chịu đựng khô hạn. Acid abscisic được
tạo ra từ lá trưởng thành và được
vận chuyển qua mô libe.
Một
trái bị thối, thì nhiều trái sẽ thối theo.
Ngày nay, người ta biết rằng trái đó
ảnh hưởng lên các trái khác là do trái thối
sinh ra khí ethylen (C2 H4). Ethylen
giữ một số vai trò trong chu kỳ đời
sống của cây, bao gồm sự tăng trưởng,
phát triển và sự lão hóa hay sự chín của trái.
Ethylen là một chất khí nên thoát ra trong không khí.
Ethylen được tổng hợp trong các mô
đang xảy ra sự lão hóa hay sắp chín như
ở trái.
Một
trong những nghiên cứu về ảnh hưởng
của ethylen là kích thích sự chín của trái.
Một trái khi đạt được một kích
thước tối đa, nó trở nên nhạy cảm
với ethylen, và đây là một hóa chất gây ra
sự chín. Quá trình chín
bắt đầu khi có sự bộc phát về các
hoạt động biến dưỡng do nồng độ
ethylen tăng lên khoảng 100 lần.
Ngăn cản sự tạo ethylen hay loại
bỏ được ethylen ngay khi chúng được
sinh ra thì ngăn chận được sự chín.
Trái được thu hoạch và vận chuyển
lúc còn xanh và cứng để tránh dập, sau đó
trái được xử lý với ethylen cho chín trước
khi đem đi bán.
Ethylen cũng thúc đẩy sự rụng của
lá, hoa và trái và góp phần vào những biến đổi
tiêu biểu sự lão hóa trong cây hoặc một
phần của cây. Hơn
nữa, ethylen còn có thể giúp phá vỡ sự miên
trạng của chồi và hột của một số
loài và gây trổ hoa ở một số loài thực
vật khác như Khóm.
|
|
Ðể
kết luận hormon trong cơ thể thực vật
điều hòa các quá trình chính trong cơ thể như
sau:
-
Tác động trên sự phân cắt của
tế bào, khâu đầu tiên của quá trình tăng trưởng
được kích thích bởi gibberellin, cytokinin và
auxin.
-
Các hormon tác động lên sự tăng trưởng
của tế bào, là giai đoạn thứ hai của quá
trình tăng trưởng, có sự tham gia của các
chất như auxin và gibberellin thúc đẩy tế bào
dài ra.
-
Các loại hormon hiện diện trong cây với tác
động phối hợp và ảnh hưởng chuyên
biệt của nó trên từng phần của cây, giúp
điều phối sự phát triển về cả
về hình thái và chức năng của cây là do tác
động kết hợp của auxin và cytokinin.
-
Qúa trình lão hóa dẫn đến sự chết,
do sự tác động của những chất gây lão
hóa khác nhau như acid abscisic, ethylen; trong đó ethylen là
một trong những chất quan trọng nhất.
III.
CÁC BẰNG CHỨNG VỀ TÍNH TOÀN NĂNG CỦA
TẾ BÀO THỰC VẬT
Số
phận của các tế bào sẽ được xác
định trong suốt quá trình phát triển.
Khi sự phát triển đã đạt được
đến một số giai đoạn nào đó thì
khi đó tế bào chỉ có thể tiến triển
theo một lộ trình duy nhất mà thôi.
Thí dụ, khối sơ khởi của lá thì
chỉ có thể phát triển thành lá mà thôi.
Tuy
vậy, ngược lại với tế bào động
vật, tế bào thực vật có tính toàn năng
(totipotency). Một
tế bào đơn độc có thể phát triển
đầy đủ và tạo ra một cơ thể hoàn
chỉnh. Ðiều này
đã được chứng minh từ 1958 bởi các
thí nghiệm của F. C. Steward (Ðại học Cornell) trên
các tế bào Cà rốt được nuôi cấy trong
môi trường vô trùng. Với
những lát cắt ngang của rễ Cà rốt, sau
đó cắt ra những miếng thật nhỏ trong vùng
mô libe thứ cấp. Các
miếng này được nuôi cấy trong môi trường
có chứa chất dinh dưỡng và chất khoáng trong
một bình tam giác và được lắc, môi trường
được thêm nước dừa tươi để
kích thích sự tăng trưởng.
Các tế bào tự do sẽ được tách ra
từ miếng mô và phân cắt; tạo ra một
cấu trúc tương tự như phôi.
Các phôi này có thể được chuyển ra và
nuôi trong môi trường đặc và dưới điều
kiện có ánh sáng chúng tăng trưởng cho ra cây Cà
rốt mới.
 |
Các mảnh nhỏ có thể được cắt ra từ thân của cây Thuốc lá và được nuôi cấy trong ống nghiệm (in vitro). Trong vài tuần, một khối mô sẹo (callus) xuất hiện từ chỗ vết cắt. Những tế bào trong mô sẹo này có thể được nuôi cấy và lưu trử để làm nguồn cung cấp tế bào. Folke Skoog (Ðại học Wisconsin) đã bố trí thí nghiệm và chứng minh rằng những tế bào trong mô sẹo cũng có tính toàn năng, có thể chuyên hóa theo những lộ trình khác nhau trong những môi trường nuôi cấy khác nhau. Tỉ lệ giữa các hormon hiện diện trong môi trường nuôi cấy cũng như nồng độ của chúng là yếu tố quyết định (Hình 15). Khi nồng độ của các hormon quá thấp thì sự tăng trưởng và chuyên hóa không xảy ra. Tuy nhiên, khi nồng độ auxin cao, rễ xuất hiện trên khối tế bào của mô sẹo: chương trình rễ (root program). Khi nồng độ cytokinin cao và auxin thấp thân được tạo ra: chương trình thân (shoot program). Với nồng độ vừa phải, tương đương nhau của cả hai hormon thì mô sẹo tiếp tục tăng trưởng và không có sự chuyên hóa: chương trình mô sẹo (callus program).
Những
lớp tế bào biểu bì được lột ra
từ những thân của cây Thuốc lá có thể
được nuôi cấy và tế bào có thể
được đi vào các chương trình chuyên hóa
(differentiating program) khác nhau.
Thí dụ, có thể tạo ra chồi dinh dưỡng,
rễ hay mô sẹo...
Từ
các thí nghiệm trên, kết luận được
rằng một tế bào thực vật đơn độc
nếu được nuôi cấy trong môi trường
dinh dưỡng và ánh sáng thích hợp có thể
trải qua sự phân cắt tế bào và tăng trưởng
để phát triển thành một cơ thể hoàn
chỉnh.
|
|
·
Kỷ
thuật nuôi cấy tế bào được sử
dụng theo phương pháp nuôi cấy vi sinh vật,
đặc biệt với các yếu tố thích hợp
cho tế bào thực vật.
Kỷ thuật nuôi cấy này là một giai đoạn
chính trong kỷ thuật di truyền để chuyển
gen. Trong công tác
chọn giống, người ta có thể nuôi cấy
tế bào đơn và tế bào trần (protoplast).
Tế bào đơn là những tế bào riêng rẻ
được tách ra từ các mô, các tế bào này có
thể được nuôi trong môi trường dinh dưỡng
và được lắc liên tục để các
tế bào không thể liên kết lại với nhau,
dung dịch lúc bấy giờ được gọi là
dịch treo tế bào (cell suspension), khi cần thiết các
tế bào này sẽ được đưa vào môi trường
thích hợp để tiếp tục nuôi cấy.
Tế bào trần là những tế bào không có vách
celluloz do được xử lý bằng hổn hợp
enzim cellulaz và macerozim. Các
tế bào này dễ dàng tiếp nhận các phân tử
ADN, plasmid và cả vi khuẩn.
Hai tế bào trần có thể được dung
hợp (fusion) với nhau để tạo ra một
tế bào lai dinh dưỡng (somatic hybridization).
· Kỷ thuật nuôi cấy mô là nuôi cấy trực tiếp từ một cấu trúc có tổ chức đầy đủ như mô phân sinh ngọn, mô phân sinh bên hay nhánh non. Kỷ thuật này được áp dụng trong vi nhân giống (micropropagation). Nuôi cấy mô phân sinh phải được khử trùng nhiệt để loại siêu khuẩn ra khỏi vùng mô này. Sự gia tăng nhiệt độ trong môi trường nuôi cấy kích thích tế bào phân cắt nhanh hơn là tốc độ phân cắt của siêu khuẩn, nhờ thế loại bỏ được siêu khuẩn ra khỏi mô. Quá trình này đã được sử dụng rộng rải để sản xuất Dâu tây và Khoai tây không có bệnh siêu khuẩn. Tuy nhiên, nuôi cấy mô từ tế bào mô sẹo thường dẫn đến kết qủa là có sự gia tăng số lượng nhiễm sắc thể hay làm thay đổi sự biểu hiện của gen hoặc gây ra đột biến. Cây được tạo ra từ nuôi cấy mô sẹo thường có cấu trúc và sinh lý không bình thường. Nên hiện nay phương pháp nhân giống từ mô sẹo thường được tránh và một kỷ thuật mới được sử dụng là nhân giống trực tiếp từ phôi soma (somatic embryo).
|
|
a.
Cứu phôi
|
|

Sự
lai giữa các loài thường tạo ra cây lai có ưu
thế do sự tổ hợp di truyền, nhưng cây
lai thường tạo ra hột không sống được
lâu. Sự nuôi cấy
in vitro để phát triển phôi từ noãn đã
thụ tinh của cây lai được dùng theo phương
pháp gọi là sự cứu
phôi lai (hay nói tắt là cứu phôi).
Thí dụ, lai giữa hai loài Cỏ ba lá Trifolium
ambiguum và Trifolium repens
thuộc họ Ðậu (Fabaceae) đã được
sử dụng để tạo ra một nguồn
giống cỏ mới cho đồng cỏ ở New
Zealand. Loài đậu
cha mẹ là các cây tự thụ phấn; phôi bình thường
phát triển từ hình cầu, hình tim đến hình
trái ngư lôi khi trưởng thành.
Tuy nhiên, phôi từ cây lai giữa hai loài trong hai ngày
đầu sau khi thụ phấn chúng tăng trưởng
nhanh, nhưng sau đó chậm lại và dừng lại
ở giai đoạn hình tim khi phôi có khoảng 1.000
tế bào. Quá trình phát
triển của phôi bị ngưng lại là do các gen gây
hại hoạt động, chúng hoạt động như
những cái khóa ngăn chận sự cung cấp
chất dinh dưỡng đến để nuôi phôi.
Một số kỷ thuật cứu phôi đã
sử dụng thành công. Ghép
phôi lai vào phôi nhũ bình thường đã được
lấy phôi ra. Phôi cũng
có thể được cấy trực tiếp trên môi
trường nhân tạo với các chất dinh dưỡng
được cung cấp theo yêu cầu.
Kỷ thuật cứu phôi này đã tạo
được cây cỏ Ba lá lai mới (Hình 16).
b.
Phôi soma
Phôi
soma là phôi được phát triển từ những
tế bào dinh dưỡng. Trong
kỷ thuật nuôi cấy phôi soma như ở Cà
rốt, thường các dung dịch treo tế bào có
thể được lưu giữ một cách không
chuyên biệt trong môi trường có chứa chất tăng
trưởng thực vật acid
2,4-dichloro-phenoxy acetic (2,4-D). Nếu
chất 2,4-D bị lấy đi sẽ cảm ứng
sự tạo phôi; phôi ở đây là phôi soma, chúng
được tạo ra từ những tế bào
của cơ thể thực vật, nhưng chúng hành
động như một hợp tử, trải qua
một quá trình phát triển như một cấu trúc
phôi. Phôi soma cũng có
thể được sản sinh từ tế bào
biểu bì của trục hạ diệp của hột
khi nuôi cấy trong môi trường có hormon; đặc
biệt là cytokinin.
c.
Nuôi cấy túi phấn và hạt phấn
Trong
các kỷ thuật nuôi cấy kể trên mô hay tế bào
đem đi nuôi cấy đều là tế bào lưỡng
bội do đó trong qúa trình
phát triển phôi sẽ xảy ra các đột biến
không mong muốn và khó có thể tạo ra các dòng
thuần. Trong kỷ
thuật nuôi cấy từ tế bào hạt phấn,
là những tế bào đơn bội nên nếu
muốn tạo ra các dòng đồng hợp tử
về một tính trạng nào đó sẽ dễ dàng hơn.
Từ những năm 20, người ta đã
chứng minh rằng các dòng lưỡng bội đồng
hợp tử có thể thu được bằng cách lưỡng
bội hóa các thể đơn bội.
Do không có tính trội lặn ở thể
đơn bội nên kiểu hình của cây phản ánh
khá trung thực kiểu gen.
Viện
Sinh vật Việt Nam đã tạo thành công cây
đơn bội từ túi phấn Thuốc lá và Lúa.
Túi phấn nuôi cấy thường được
chọn ở giai đoạn hạt phấn đơn
nhân trước khi nhân hạt phấn phân chia để
tạo ra hai nhân dinh dưỡng và nhân sinh dục.
Trong môi trường nuôi cấy thích hợp
tế bào hạt phấn đơn nhân phân chia lần
đầu tạo ra hai nhân và sau đó tiếp tục
phân chia để tạo ra tiền phôi.
Trong các kết quả nuôi cấy có thể thu
được những cây đa bội tự nhiên bên
cạnh các cây đơn bội.
Nếu muốn lưỡng bội hóa các cây
đơn bội có thể xử
lý bằng colchicin, các cây con lưỡng bội này
được gọi là cây đơn bội kép
(dihaploid).
![]()
![]()