CHƯƠNG
1
CẤU TRÚC CỦA TẾ BÀO
Cơ thể sinh vật được cấu
tạo bởi những đơn vị cơ bản
được gọi là tế bào.
Cơ thể con người được cấu
tạo bởi hàng ngàn tỉ tế bào.
Trẻ sơ sinh có khoảng 2.000 tỉ tế bào,
người trưởng thành có khoảng 100.000 tỉ
tế bào; có khoảng 30 tỉ tế bào trong não, 20
tỉ tế bào hồng cầu trong máu và có khoảng
200 loại tế bào chuyên hóa khác nhau.
Trong khi đó vi khuẩn và các vi sinh vật, cơ
thể chỉ là một tế bào.
Hầu hết tế bào không thấy được
bằng mắt trần nên những hiểu biết
về tế bào tùy thuộc vào trình độ phát
triển của kính hiển vi.
Vào các năm 50, các nhà sinh vật học mới
biết có 5 hay 6 bào quan hiện diện bên trong tế bào,
nhưng hiện nay với kính hiển vi điện
tử người ta đã quan sát được ở
mức siêu cơ cấu của rất nhiều bào quan
hiện diện trong tế bào.
I.
ÐẠI CƯƠNG VỀ TẾ BÀO
1. Lược sử phát hiện tế bào |
|
Hầu
hết các tế bào đều có kích thước
rất nhỏ nên mắt trần không thể quan sát
được, do đó lược sử phát hiện
tế bào gần như là lịch sử phát minh ra kính
hiển vi. Galileo (1564 -
1642) chế tạo ra viễn vọng kính để quan
sát bầu trời, tình cờ khám phá ra những
vật rất nhỏ khi quan sát bằng cách lật ngược
đầu kính lại.
Antoni Van Leeuwenhoek (1632 - 1723) người Hà Lan, do yêu
cầu kiểm tra tơ lụa, ông mài các thấu kính
để quan sát chất lượng của vải,
nhờ đó quan sát được những vật li
ti quanh môi trường sống và khám phá ra sự
hiện diện của thế giới vi sinh vật.
Robert
Hooke (1635 - 1703) người Anh, lần đầu tiên mô
tả các lỗ nhỏ có vách bao bọc của
miếng bấc (nút bần) cắt ngang dưới kính
hiển vi năm 1665 và Hooke dùng thuật ngữ tế bào
(cellula có nghĩa là phòng, buồng nhỏ, vì ý nghĩa
lịch sử từ này vẫn còn được dùng
cho đến ngày nay) để chỉ các lỗ đó.
2. Thuyết tế bào |
|
Mãi đến thế kỷ 19 khái niệm sinh vật có
cấu tạo tế bào của Hooke mới được
sống dậy từ nhiều công trình nghiên cứu,
đặc biệt hai công trình của hai người Ðức:
nhà thực vật học Matthias Jakob Schleiden (1838) và nhà
động vật học Theodor Schwann (1839).
Hai ông đã hệ thống hóa quan điểm thành
thuyết tế bào Tất cả các
sinh vật do một hay nhiều tế bào tạo thành,
nói một cách khác, Tế bào là đơn vị
cấu tạo sống cơ bản của tất
cả sinh vật. Ðến
năm 1858 thuyết tế bào được mở
rộng thêm do một bác sĩ người Ðức
(Rudolph Virchow): Tế bào do tế bào có trước sinh
ra. Quan điểm
(mở rộng tế bào) của Virchow sau đó
được Louis Pasteur (1862) thuyết phục các nhà
khoa học đồng thời bằng hàng loạt thí
nghiệm chứng minh. Như
vậy có thể tóm tắt thuyết tế bào như
sau: Tế bào là đơn vị cấu tạo sống
cơ bản của tất
cả sinh vật, tế bào do tế bào có trước
sinh ra.
3. Hình dạng và kích thước tế bào |
a.
Hình dạng
Hình
dạng của tế bào rất biến thiên và tùy
thuộc rất nhiều vào tế bào là một sinh
vật đơn bào hay tế bào đã chuyên hóa để
giữ một nhiệm vụ nào đó trong cơ
thể sinh vật đa bào.
Từ những dạng đơn giản như hình
cầu, hình trứng, hình que có thể gặp ở các
sinh vật đơn bào đến những hình
dạng phức tạp như các tế bào hình sao
ở mô thực vật, hay các tế bào thần kinh
ở động vật cấp cao...
Ðặc
biệt ở các sinh vật đơn bào hình dạng có
ý nghĩa quan trọng đối với đời
sống của chúng. Thí
dụ, vi khuẩn hình cầu có thể chịu đựng
được sự khô hạn giỏi vì diện tích
tiếp xúc với môi trường bên ngoài ít do đó
giữ được nước dù môi trường
sống rất khô. Ngược
lại vi khuẩn hình que dài có diện tích tiếp xúc
cho mỗi đơn vị thể tích với môi trường
bên ngoài lớn hơn nên có thể tồn tại
dễ dàng trong môi trường có nồng độ
thức ăn không cao.
b.
Kích thước
Kích
thước của tế bào cũng rất biến thiên
theo loại tế bào. Nói
chung, thường tế bào rất nhỏ và phải dùng
kính hiển vi mới quan sát được.
Vi khuẩn có lẻ là những sinh vật
đơn bào có kích thước nhỏ nhất.
Thí dụ, vi khuẩn Dialister pneumosintes có kích thước
rất nhỏ 0,5 x 0,5 x 1,5 (m trong khi trứng của chim
đà điểu là tế bào có đường kính
đến 20 cm, hay tế bào thần kinh có đường
kính nhỏ nhưng có thể dài đến 90 - 120 cm.
Trung bình thì đường kính biến thiên trong
khoảng từ 0,5 đến 40 (m).
Thật ra độ lớn nhỏ của tế bào không quan trọng mà tỉ lệ giữa diện tích bề mặt và thể tích tế bào mới có ảnh hưởng lớn đến đời sống của một tế bào. Tế bào lấy thức ăn, oxy từ môi trường chung quanh và thải chất cặn bả ra bên ngoài tế bào. Các vật liệu này đều phải di chuyển xuyên qua bề mặt của tế bào. Khi tế bào gia tăng kích thước, thể tích tăng gấp nhiều lần so với sự gia tăng của diện tích (ở hình cầu, thể tích tăng theo lủy thừa bậc ba trong khi diện tích tăng theo lủy thừa bậc hai). Do đó, khi tế bào càng lớn lên thì sự trao đổi qua bề mặt tế bào càng khó khăn hơn
4. Phân loại tế bào |
Dựa
trên đặc điểm cấu trúc của tế bào
có thể phân chia tế bào của sinh vật ra làm hai
nhóm: tế bào sơ hạch và tế bào chân hạch.
*
Tế bào sơ hạch là loại tế bào không có màng
nhân. ADN có kiến trúc
xoắn vòng kín. Không có
các bào quan có màng. Các
tế bào này gặp ở các sinh vật thuộc
giới sinh vật sơ hạch: Archaebacteria và
Eubacteria.
*
Tế bào chân hạch là loại tế bào có nhân
với màng nhân bao quanh, và nhiều loại bào quan có màng
bao. Các tế bào này
gặp ở các sinh vật thuộc các giới Protista,
Nấm, Thực vật và Ðộng vật.
II.
CẤU TRÚC CỦA TẾ BÀO CHÂN HẠCH (Hình 1 và 2)
1. Màng tế bào |
Tế
bào được một màng bao bọc gọi là màng
tế bào, bên trong màng là chất
nguyên sinh (protoplasm), gồm tế bào chất (cytoplasm),
nhân và các bào quan (organelle) khác (Cấu trúc của màng
sẽ được đề cập trong chương
2).
2. Các bào quan |
|
a.
Mạng nội chất (endoplasmic reticulum: ER)
Mạng
nội chất được phát hiện vào năm
1945 dưới kính hiển vi tương phản pha
(phase-contrast microscope). Mạng
nội chất hiện diện ở tất cả
tế bào chân hạch. Kính
hiển vi điện tử cho thấy mạng nội
chất nối liền với màng ngoài của nhân
ở một số vị trí.
Hình 3a. Mạng
nội chất sần
Hình 3b. Mạng
nội chất láng
Vùng ngoại vi của màng nhân liên tục với
khoang của mạng nội chất.
Do đó, những
kênh trên mạng nội chất có thể là con
đường để vận chuyển vật
chất giữa nhân và những phần khác của
tế bào chất, tạo ra một hệ thống thông
tin giữa nhân là trung tâm điều khiển và
phần còn lại của tế bào.
Hầu hết protein liên kết với màng hay
được vận chuyển bởi mạng nội
chất được tổng hợp bởi ribô
thể của mạng nội chất sần.
Protein tổng hợp từ các ribô thể tự
do trong tế bào chất sẽ thực hiện chức
năng trong dịch tế bào chất.
Nhiệm vụ của mạng nội chất không
đơn thuần là đường vận chuyển bên
trong tế bào. Màng
của mạng nội chất là nơi chứa các
protein và các protein này có cả hai chức năng,
vừa là thành phần cấu trúc vừa là enzim xúc tác
các phản ứng hóa học.
Hiện nay, có nhiều bằng chứng cho thấy
rằng ít nhất là một số protein cấu tạo
mạng nội chất hoạt động như enzim;
một số của những enzim này được
gắn thêm một đường đa ngắn,
đường này có nhiệm vụ như một cái
nhản đưa thư
(mailing label) để đưa protein đến
đúng nơi nhận trong tế bào. Trong hình 4 một
chuỗi gồm 14 phân tử đường gắn vào
phân tử protein trên
mạng nội chất sần có chức năng như
một cái nhản, protein nào không có nhản thì sẽ
ở lại mạng nội chất.
Khi bốn phân tử đường cuối cùng
được tách ra thì protein sẽ được
chuyển đến hệ Golgi (G: glucoz, M: manoz, N:
N-acetyl-glucosamin).
Mạng nội chất còn có
nhiệm vụ như một xưởng chế tạo, các enzim của chúng xúc tác sự tổng hợp các
phospholipid và cholesterol được dùng để tạo ra màng mới hay các protein
màng được tổng hợp bởi ribô thể trên mạng nội chất là thành phần của màng
lipid mới. Vùng láng đặc biệt
gia tăng ở các tế bào có nhiệm vụ tổng hợp lipid như hormon steroid.
Ở tế bào gan của động vật có xương sống, protein màng của vùng láng
có vai trò quyết định trong sự thải chất độc và dược phẩm như các thuốc giảm
đau (barbiturate), các loại thuốc kích thích như camphetamin, morphin và
codein. Hình
4. Protein được
gắn nhản Hệ
Golgi (do Camillo Golgi, người đầu tiên mô tả
vào năm 1898) gồm một hệ thống túi dẹp
có màng bao và xếp gần như song song nhau (Hình 5).
Mặt phía gần nhân được gọi là
mặt cis, phía đối diện là mặt trans. Các túi
chuyên chở (transport vesicle) chứa bên trong lipid và
protein mới được tổng hợp, được
tách ra từ màng của mạng nội chất hòa vào
các túi dẹp của hệ Golgi ở mặt cis.
Các chất này vào trong hệ Golgi được
biến đổi, sắp xếp lại và sau đó các
túi mới được tách ra từ mặt trans. Các
túi này vận chuyển các phần tử đến các
bào quan khác và màng sinh chất, đôi khi các túi
được chuyển đến glycocalyx.
Hệ Golgi đặc biệt to ở những tế bào
tiết như tế bào tụy tạng tiết ra insulin
hay tế bào ruột
non tiết ra chất nhày (mucus).
Hiện nay, người ta biết vai trò của
hệ Golgi trong chức năng tiết là tồn
trử, biến đổi (cô đặc lại) và
bọc các sản phẩm tiết lại.
Mặc dù sự tổng hợp protein không xảy
ra ở hệ Golgi, nhưng những đường
đa được tổng hợp tại đây
từ các đường đơn sẽ được
gắn vào lipid hay protein để tạo ra glycolipid hay
glycoprotein. Các túi
được tách ra từ hệ Golgi có vai trò
quan trọng làm tăng bề mặt của màng
tế bào. Khi túi
được chuyển đến bề mặt
của màng sinh chất, chúng sẽ được
gắn vào màng này, sau đó vỡ ra và phóng thích
nội dung ra bên ngoài tế bào trong quá trình ngoại
xuất bào, một phần hay tất cả màng của
túi được hòa vào màng sinh chất hay trở
về hệ Golgi.
Các kết quả nghiên cứu cho thấy có
sự liên quan giữa những phần khác nhau của
tổng thể hệ thống màng trong tế bào.
Một phân tử cấu trúc màng có nguồn
gốc từ vùng sần của mạng nội chất
được chuyển đến vùng láng, sau đó
đi đến hệ Golgi trong các túi chuyên chở
và cuối cùng đến màng sinh chất, từ
đây chúng có thể trở về hệ Golgi hay
một số bào quan khác chỉ là một túi rổng.
Như vậy màng phospholipid luôn được
đổi mới. Tiêu
thể là một thể có màng bao bọc, là những túi
dự trử các enzim có khả năng thủy phân các
đại phân tử trong tế bào.
Màng của tiêu thể là màng không thấm, bên
trong chứa các enzim tiêu hóa.
Nếu màng của tiêu thể bị vỡ ra, các
enzim được phóng thích vào trong tế bào chất
và lập tức các đại phân tử trong tế bào
sẽ bị thủy giải.
Tiêu thể hoạt động như một
hệ thống tiêu hóa của tế bào, có khả năng
tiêu hóa các vật liệu có kích thước lớn
được mang vào tế bào do sự nội
nhập bào.Trong hình 6 enzim tiêu hóa được
tổng hợp ở vùng sần của mạng nội
chất, được bọc lại ở vùng láng
trong các túi chuyên chở và được chuyển
đến hệ Golgi.
Khi có một nơi nào đó cần enzim thì
từ màng của hệ Golgi tách ra một túi có
chứa enzim. Những
protein gắn bên ngoài của tiêu thể như là
một bộ phận để nhận diện ra
rằng enzim đã đến đúng vị trí cần
nó, thí dụ nơi có các túi nội nhập bào từ
màng tế bào đưa vào, tại đây tiêu thể
và túi này sẽ hòa vào nhau.
Khi nội dung được tiêu hóa hoàn toàn,
những sản phẩm hữu ích được
đưa trở vào tế bào chất, những cặn
bả được đưa ra ngoài bởi sự
ngoại xuất bào, màng của túi được hòa
nhập vào màng tế bào.
Sự hoạt động không bình thường
của tiêu thể là nguyên nhân của nhiều
bệnh. Một trường hợp đã được
biết là bệnh Tay - Sachs, vì tiêu thể tiêu hóa lipid
thiếu một enzim. Khi
những tiêu thể thiếu enzim này hòa vào những túi
chứa lipid, chúng không tiêu hóa hoàn toàn được
nội dung này. Kết
quả là những túi này tích tụ và làm nghẻn
sự dẫn truyền các xung động của các
tế bào thần kinh. d.
Peroxisom Peroxisom
có hình dạng tương tự tiêu thể, nhưng
peroxisom không được tách ra từ hệ Golgi, mà
nó được sinh ra từ peroxisom có trước.
Tế bào phải được thừa hưởng
ít nhất một
peroxisom từ tế bào chất của tế bào
mẹ, nếu không sẽ không tránh khỏi cái
chết.
Tương tự như tiêu thể, peroxisom
chứa enzim nhưng là enzim oxy hóa, chúng xúc tác các
phản ứng trong đó nguyên tử hydro được
chuyển từ hợp chất hữu cơ (như
formaldehyd và rượu ethyl) đến oxy, để
tạo ra hydro peroxyd (H2O2), đây là một chất
cực độc đối với tế bào.
Tuy nhiên, peroxisom còn có một enzim khác nữa là
catalaz, sẽ chuyển chất H2O2 độc này thành nước
và oxy. Tế bào gan và
thận người có rất nhiều peroxisom, do đó
rượu ethyl do người uống được
oxy hóa nhờ các peroxisom trong những tế bào này. e.
Không bào (vacuole) Không
bào có một màng bao quanh, bên trong chứa một
dịch lỏng , được tìm thấy cả
ở tế bào thực vật và động vật,
đặc biệt rất phát triển ở tế bào
thực vật. Có
nhiều loại không bào với chức năng khác
nhau. Ở một số động vật nguyên sinh,
đặc biệt sống ở nước ngọt có
không bào đặc biệt gọi là không bào co bóp
(contractile vacuole) giử vai trò quan trọng trong sự
thải nước ra khỏi tế bào.
Không bào tiêu hóa để tiêu hóa thức ăn.
Ngoài ra, ở vi khuẩn và vi khuẩn lam có không bào
khí chứa khí giúp tế bào nổi lên mặt nước.
Ðiểm đặc biệt là màng bao của không
bào khí được cấu tạo bằng protein.
Ở hầu hết tế bào thực vật, có
một không bào rất to chiếm từ 30 - 90% thể tích
tế bào. Các tế bào
chưa trưởng thành có nhiều không bào nhỏ
xuất xứ từ mạng nội chất và hệ
Golgi. Các túi này tích
chứa nước, to ra và có thể hòa vào nhau để
tạo ra một không bào to ở tế bào trưởng
thành. Dần dần, không
bào sẽ đẩy tế bào chất ra ngoại biên
của tế bào và chỉ còn là một lớp
mỏng (Hình 7). Không
bào ở tế bào thực vật chứa một
dịch lỏng gồm nước và một số
chất hòa tan trong đó. Màng
không bào có tính thấm đặc biệt và có thể
điều khiển sự di chuyển vật chất
qua màng này. Không bào tích
nước bằng sự thẩm thấu, trương
lên và đẩy tế bào chất ra sát vách tế bào,
áp lực này tạo cho tế bào một sức trương
nhất định. Vách
tế bào đủ cứng rắn để giử cho
tế bào không bị vỡ ra.
Sức trương của tế bào rất quan
trọng, nó giúp cho các cơ quan của cây như lá, thân
non đứng vững được.
Khi sức trương này không còn nữa, cây
sẽ héo. Nhiều
chất quan trọng cho đời sống của
thực vật được chứa trong không bào: các
hợp chất hữu cơ
như đường, các acid hữu cơ, acid
amin, một vài protein, sắc
tố antocianin cho màu tím, xanh và đỏ đậm thường
thấy trong cánh hoa, trái và lá vào mùa thu.
Aïp suất thẩm thấu cao của không bào giúp
cho cây có thể hấp thu được nước
ở đất rất khô bằng sự thẩm
thấu. Ngoài ra, không bào
còn là nơi tích chứa những chất thảií do các
quá trình biến dưỡng.
Một số chất thải có thể được
xử dụng lại dưới tác dụng của
enzim. Chức năng này
rất quan trọng vì cây không có thận hay các cơ
quan khác để thải chất bả như động
vật, thực vật thải chất bả khi cây
rụng lá. f.
Ty thể bộ (mitochondria) Ty
thể bộ là toàn thể các ty thể hiện
diện trong tế bào. Ty
thể là nơi tổng hợp năng lượng
chủ yếu của tế bào chân hạch, là nơi
diễn ra quá trình hô hấp, lấy năng lượng
từ thức ăn để tổng hợp ATP là
nguồn năng lượng cần thiết cho các
hoạt động của tế bào.
Năng lượng cần thiết để co cơ
hay cung cấp cho các bơm hoạt động trong
sự vận chuyển tích cực qua màng tế bào.
Số lượng ty thể tùy thuộc vào
mức độ hoạt động của tế bào.
Thí dụ, tế bào cơ tim có 72 lần co
thắt trong một phút có chứa hàng ngàn ty thể.
Giống như nhân, mỗi ty thể được
bao bọc bởi hai màng, màng ngoài trơn láng, màng trong
với các túi gấp nếp (crista) sâu vào bên trong
chất căn bản (matrix) làm gia tăng diện tích
của màng trong lên rất nhiều (Hình 8).
Ty thể có chứa ADN,
ribô thể riêng nên có
thể nhân lên độc lập với sự phân chia
của nhân.
Hình 8. Cơ
cấu của ty thể g.
Lạp bộ (plastid) Lạp
bộ gồm tất cả các lạp hiện diện
trong tế bào, được tìm thấy trong hầu
hết tế bào thực vật, trừ tế bào
nấm, mốc và tế bào động vật.
Lạp có thể quan sát được dưới
kính hiển vi thường.
Có hai loại lạp chính: sắc lạp
(chromoplast) và vô sắc lạp (leucoplast).
*
Lục lạp (chloroplast) Lục
lạp là sắc lạp có chứa diệp lục
tố (chlorophyll), và các sắc tố vàng hay cam
gọi là carotenoid. Phân
tử diệp lục tố hấp thu năng lượng
ánh sáng mặt trời để tổng hợp ra các
phân tử hữu cơ phức tạp (đặc
biệt là đường) từ các nguyên liệu vô cơ
như nước và khí carbonic.
Phản ứng tổng hợp này sản sinh ra oxy,
là loại khí mà hầu hết các loài sinh vật khác
lệ thuộc vào nó.
Dưới kính hiển vi điện tử,
một lục lạp được bao bọc bởi
hai màng và vô số các
túi dẹp có màng bao được gọi là thylakoid
nằm trong chất cơ bản gần như
đồng nhất được gọi là stroma.
Thylakoid hoặc phân bố khắp trong stroma,
hoặc xếp chồng chất lên nhau được
gọi là grana (Hình 9). Diệp
lục tố và carotenoid gắn trên màng thylakoid.
Lục lạp cũng có chứa ADN và ribô thể
riêng như ty thể. *
Sắc lạp không có diệp lục tố Thường
có màu vàng hay cam (đôi khi có màu đỏ) vì chúng
chứa carotenoid. Sắc
lạp này làm cho hoa, trái chín, lá vàng có màu vàng hay cam
đặc trưng. Một
số sắc lạp không bao giờ chứa diệp
lục tố, một số khác thì do mất diệp
lục tố, đây là trường hợp của trái
chín và lá mùa thu. *
Vô sắc lạp Vô
sắc lạp có chứa các vật liệu như tinh
bột, dầu và protein dự trử.
Lạp có chứa tinh bột được
gọi là bột lạp (amyloplast), thường gặp
ở hột như lúa và bắp, hay dự
trử trong rễ và thân như carot và khoai tây, ngoài
ra có thể hiện diện trong tế bào ở các
phần khác của cây. Tinh
bột là hợp chất dự trử năng lượng
dưới dạng từng hạt.
Cây có hột giàu tinh bột là nguồn lương
thực giàu năng lượng.


![]()
b.
Hệ Golgi

Hình 5.
Hệ Golgi
c.
Tiêu thể (lysosome)

Hình 6.
Chu trình của enzim tiêu hóa![]()

Hình
7. Sự phát triển
của không bào ở tế bào thực vật


Hình 9.
Cơ cấu của lục lạp (dưới kính
hiển vi điện tử)

Hình
10. Ribô thể
![]()
h.
Ribô thể
Tế bào chất chứa vài triệu ribô thể,
ribô thể là những hạt nhỏ không có màng bao có
đường kính từ 25 - 30 nm.
Ribô thể gồm hai bán đơn vị
được tạo ra trong hạch nhân từ
những phân tử ARN và protein (Hình 10).
Ở tế bào chân hạch các bán đơn
vị này đi qua lỗ của màng nhân ra ngoài tế
bào chất, nơi đây chúng sẽ kết hợp
với phân tử mARN để tổng hợp protein.
Ribô thể trược dọc theo sợi mARN
tạo ra một chuổi ribô thể được
gọi là polyribosom hay
polysom, các ribô thể sau khi tổng hợp protein vẫn
tiếp tục tự do trong tế bào chất hay chúng có
thể gắn vào bề mặt của mạng nội
chất. Các ribô
thể có thể gắn trên vùng sần của mạng
nội chất hay trôi nổi trong tế bào chất.
Protein được tổng hợp từ các ribô
thể tự do trong tế bào chất thì không
được đưa ra khỏi tế bào hay tham gia
vào cấu trúc màng tế bào mà là những enzim trong
dịch tế bào chất.
i.
Trung thể (centrosome)
Ở hầu hết tế bào động vật, bên ngoài nhân có một vùng được gọi là trung thể bao gồm hai bào quan được gọi là trung tử (centriole). Trung tử hiện diện từng đôi và xếp thẳng góc nhau. Khi có trung tư,í chúng là nơi xuất phát của thoi vi ống trong lúc tế bào phân cắt. Ở tiết diện ngang, trung tử có cấu trúc đồng đều với chín nhóm ba vi ống xếp thành đường tròn (Hình 11).
Thể
gốc (basal bodies) có cấu tạo giống hệt trung
tử, được tìm thấy ở gốc của các
tiêm mao và
chiên mao. Trung tử và
thể gốc có mối liên quan với nhau.
Thí dụ, thể gốc ở chiên mao của
nhiều loại tinh trùng
trở thành một trung tử của tế bào
trứng sau khi thụ tinh; trung tử của tế bào
ở thành của ống dẫn trứng phân cắt thành
thể gốc tiêm mao hoạt động nhịp nhàng
để đưa trứng đến nơi thụ
tinh.
|
|

Hình
11. Tiết diện ngang
trung tử
3. Nhân (nucleus) |
Nhân là bào quan lớn nhất có màng bao và quan sát được rõ ràng nhất trong các tế bào chân hạch. Nhân đóng vai trò quan trọng trong sự sinh sản của tế bào. Nhân không những là trung tâm của mọi hoạt động của tế bào mà nó còn có vai trò quan trọng trong việc xác định các đặc điểm của thế hệ con cháu của chúng. Nhân chứa hai cấu trúc phân biệt được là nhiễm sắc thể (chromosome) và hạch nhân (nucleolus). Dưới kính hiển vi điện tử, có thể thấy được hai cấu trúc này nằm trong một khối chất vô định hình có dạng hạt, được gọi là chất nhân (nucleoplasm). Nhân được bao bọc bởi màng nhân (nuclear envelope) gồm hai màng phân biệt được.
a.
Nhiễm sắc thể
 |
 |
||
Hình 12. Sơ đồ thể nhân Hình 13. Màng nhân
Nhiễm
sắc thể hình sợi dài chỉ quan sát được
rõ ràng trong lúc tế bào đang phân chia mà thôi, gồm
ADN và protein. ADN là
vật liệu chứa các đơn vị cơ
bản của sự di truyền được gọi
là gen (gene), protein làm thành những phần lõi giống
như những cuộn chỉ, sợi ADN quấn lên
đó, để thành lập cấu trúc thể nhân
(nucleosome) (Hình 12). Gen
được sao chép khi tế bào phân cắt để
mỗi tế bào con đều có một bản sao.
Tất cả gen trong tế bào được
gọi là bộ gen (genom).
Thông tin di truyền mang bởi các gen là trình
tự các nucleotid, của phân tử ADN.
Trình tự này xác định trình tự của
acid amin trong phân tử protein được tổng
hợp trong tế bào chất.
Do vậy, gen được xem là trung tâm của
sự sống, chúng mã hóa các thông tin cần thiết
cho sự tổng hợp các enzim, để điều
hòa vô số các phản ứng hóa học tiêu biểu
của tế bào và của sinh vật.
b.
Hạch nhân
Hạch
nhân thường được thấy rõ nhất lúc
tế bào không phân chia. Nhân chứa một đến
nhiều hạch nhân. Là
một phần của nhiễm sắc thể nên
hạch nhân cũng gồm ADN và protein.
ADN của hạch nhân gồm nhiều bản sao
của gen làm khuôn tổng hợp rARN cho ribô thể.
Sau khi được tổng hợp, rARN kết
hợp với protein rồi rời khỏi hạch nhân
và đi ra tế bào chất, nơi đây chúng trở
thành một thành phần của ribô thể.
Do vậy, khi tế bào ít tổng hợp protein
hạch nhân rất nhỏ hay gần như vắng
mặt.
c.
Màng nhân
Màng
nhân ngăn cách môi trường bên trong nhân và tế bào
chất bao quanh. Màng nhân
còn là nơi cho hai đầu nhiểm sắc thể bám
vào. Không giống như
màng sinh chất, màng nhân gồm hai màng, màng ngoài và màng
trong, khoảng ngăn cách giữa hai màng là vùng
ngoại vi. Dưới
kính hiển vi điện tử hai màng của màng nhân
được ngắt quảng bởi các lỗ,
mỗi lỗ nhân được viền bởi một
phức hợp gồm tám protein (Hình 13).
Màng nhân có tinh thấm chọn lọc cao.
Sự trao đổi chất xuyên qua các lỗ
được kiểm soát nghiêm ngặt và có tính
chọn lọc rất cao. Nhiều
nghiên cứu cho thấy một số phân tử có
thể đi xuyên qua màng sinh chất vào tế bào
chất, nhưng lại không thể xuyên qua màng nhân
để vào nhân, ngay cả những phân tử có kích
thước nhỏ hơn lỗ nhân; trong khi đó có
những phân tử lớn hơn lỗ nhân lại có
thể đi qua được.
4. Tế bào chất |
|
Tế
bào chất là thành phần của chất nguyên sinh
gồm phần dịch lỏng là dịch tế bào
chất (cytosol) và bộ khung protein được
gọi là khung xương tế bào (cytoskeleton).
a.
Dịch tế bào chất
Là
phần dịch lỏng của tế bào chất, không
màu, hơi trong suốt, đàn hồi, không tan trong nước.
Thành phần chính của dịch tế bào
chất là nước, ngoài ra còn có các đại phân
tử protid, lipid và glucid. Các
đại phân tử này làm thành các micel, các micel mang cùng
điện tích đẩy nhau gây nên chuyển động
Brown (Robert Brown) của tế bào chất.
Do sự hiện diện của các micel nên
dịch tế bào chất ở trạng thái keo, vừa
có thể ở trạng thái lỏng (sol) vừa ở
trạng thái đặc (gel).
Trong tế bào sống thường xuyên có sự
thay đổi giữa hai trạng thái trên.
Dịch tế bào chất là nơi xảy ra các
phản ứng của các quá trình biến dưỡng
trong tế bào, các quá trình này sẽ được
đề cập trong các chương sau.
b.
Khung xương tế bào
Các bào quan trôi nổi tự do trong dịch tế bào chất của tế bào chân hạch hoặc chúng có thể gắn vào hệ thống sợi protein phức tạp bên trong, được gọi là khung xương tế bào. Cái sườn protein này tạo hình dạng của tế bào, tham gia vào các cử động của tế bào và đặc biệt quan trọng trong lúc tế bào phân chia. Ba thành phần quan trọng nhất của khung xương tế bào là vi sợi (microfilament), sợi trung gian
(intermediate
filament), và vi ống (microtubule).
Ba thành phần này được cấu tạo
bởi những bán đơn vị (subunit) protein có
thể tập họp thành sợi đơn hay đôi và
có thể làm thay đổi hình dạng của tế bào
hay gây ra các cử động.* Vi sợi

Vi sợi dài,
cực mảnh, làm thành sợi đôi, quấn xoắn
do các protein actin trùng hợp tạo thành (Hình 14).
Vi sợi actin giử vai trò cấu trúc, chúng đan
chéo nhau giữ hình dạng tê úbào.
Sợi actin khi kết hợp với myosin tham gia vào
sự cử động của tế bào.
Sợi myosin dài, mãnh, rất giống sợi actin,
nhưng có một đầu to.
Ðiểm đặc trưng của sự kết
hợp actin-myosin là khi được cung cấp năng
lượng ATP thì phần đầu của sợi
myosin móc vào sợi actin và uốn ngược lại (Hình
15). Sự cử động
này làm cho màng cử động kéo theo sự cử
động của các sợi actin khác. Ðiều này
giải thích được các chuyển động như
sự co cơ, sự vận chuyển của các túi
chuyên chở bên trong tế bào, vùng giữa của
tế bào mẹ thắt lại tách hai tế bào con,
cử động ở amip.


*
Sợi trung gian
Các vi sợi trung gian được tìm thấy nhiều ở tế bào chịu nhiều kích thích cơ học như tế bào thần kinh nên có lẻ chúng làm thành cái khung chống đỡ cho tế bào và nhân của chúng. Bán đơn vị căn bản cấu tạo nên sợi trung gian gồm hai phân tử protein quấn xoắn nhau. Bán đơn vị căn bản này xếp thành từng đôi (một tứ phân), các đôi này gắn nối tiếp nhau từ đầu đến đuôi tạo thành sợi giống như dây thừng. Tám sợi đôi này tạo thành một ống rỗng (Hình 16).
* Vi ống

Hình 17. Vi ống
Vi ống giử vai trò quan trọng trong hệ thống
mạng của khung xương tế bào.
Vi ống có cấu trúc hình trụ dài, rổng
cấu tạo bởi các phân tử protein hình cầu
tubulin. Mỗi tubulin
gồm hai protein (( và (), trùng phân thành một chồng
xoắn ốc gồm 13 bán đơn vị (Hình 17).
Vi ống tăng trưởng nhờ sự gắn
thêm vào của những phân tử tubulin vào một
đầu của sợi; vi ống giử vai trò quan
trọng trong cấu trúc và trong sự phân cắt
của tế bào. Trong
lúc tế bào phân chia vi ống được thành
lập và tỏa ra từ mỗi cực của tế bào
để tạo ra thoi vi ống để các nhiễm
sắc thể trượt trên đó về hai cực
của tế bào hình thành nhân mới.
Giống
như vi sợi, chúng cũng tham gia vào sự chuyển
vận bên trong tế bào và giúp tạo hình dạng và
nâng đỡ cho tế bào cũng như các bào quan
của nó. Vi ống còn
có vai trò chính trong cấu trúc và cử động
của tiêm mao và chiên mao.
| 5. Vách tế bào và vỏ tế bào |
|
a.
Vách tế bào thực vật
Vách
tế bào là đặc điểm của tế bào
thực vật để phân biệt với tế bào
động vật, vách bảo vệ tế bào, giữ
hình dạng, tránh sự mất nước cũng như
chống sự xâm nhập của các vi sinh vật.
Vách ở phía ngoài của màng có thể dày
từ 0,1 đến vài
(m. Thành phần hóa
học của vách thay đổi từ loài này sang loài
khác và từ tế bào này sang tế bào khác trong cùng
một cây, nhưng cấu trúc cơ bản không thay
đổi. Thành
phần cấu tạo chính là các phân tử celluloz có
dạng sợi được kết dính với nhau
bằng chất nền gồm các đường đa
khác và protein.
Các
phân tử celuloz cấu trúc thành các sợi celluloz
xếp song song nhau tạo ra các tấm, các sợi trong các
tấm khác nhau thường tạo ra các góc từ 60 -
90o. Ðặc điểm
sắp xếp này làm vách tế bào rắn chắc.
Các sợi celluloz rộng khoảng 20 nm, giữa các
sợi có những khoảng trống có thể cho nước,
khí và các ion di chuyển tự do qua mạng lưới
này, tính thấm chọn lọc của tế bào là do màng
sinh chất quy định (Hình 18).
Ở những cây còn non tế bào có vách mỏng
gọi là vách sơ cấp (primary wall), vách này có tính
đàn hồi và cho phép tế bào gia tăng kích thước.
Giữa hai vách sơ cấp của các tế bào
liên kề nhau là phiến giữa hay lớp chung (middle
lamella), là một lớp mỏng giàu chất polysaccharid
gọi là pectin, thường hiện diện dưới
dạng là pectat calci. Khi
chất pectin bị hóa nhày, các tế bào không còn
gắn chặt vào nhau nữa như khi chín trái trở
nên mềm đi.


Hình
18. Các sợi celluloz
trong
vách
tế bào (kính hiển vi điện tử)
Hình 19. Vách
của ba tế bào liên kề
Khi tế bào trưởng thành và ngừng tăng
trưởng, một số tế bào tạo thêm
lớp cứng hơn gọi là vách thứ cấp
(secondary wall) nằm giữa vách sơ cấp và màng
tế bào. Vách thứ
cấp thường dày có nhiều lớp được
cấu tạo bằng các sợi celluloz xếp theo
nhiều hướng khác nhau, nên vách tế bào trở
nên rắn chắt hơn. Ngoài
celluloz vách thứ cấp còn có thể tẩm thêm
nhiều chất khác như lignin.
Khi vách thứ cấp được thành lập
hoàn toàn, tế bào có thể chết đi, khi đó
chỉ còn làm nhiệm vụ nâng đỡ hay dẫn
truyền (Hình 19).
Trên vách của tế bào thực vật có
những lỗ nhỏ giúp các chất thông thương
giữa các tế bào với nhau, các lỗ này
được gọi là cầu liên bào (plasmodesmata),
ở vị trí này tế bào chất của hai tế bào
liên kề liên tục nhau (symplast).
b.
Vỏ của tế bào động vật (coat)
|
|

Ở thực vật, nấm và vi khuẩn, vách tế bào riêng biệt với màng tế bào, ngược lại ở tế bào động vật, vỏ không độc lập với màng. Các carbohydrate cấu tạo nên vỏ tế bào thành lập các cầu nối hóa trị với các phân tử lipid hay protein trên màng tế bào (Hình 20). Kết quả tạo ra các glycolipid hay glycoprotein, do đó vỏ tế bào còn được gọi là glycocalyx. Lớp này nằm bên ngoài Hình 20. Màng tế bào với glycocalyx của màng đôi lipid. Theo các nghiên cứu mới đây, trong glycocalyx có những điểm nhận diện (recognition site) trên bề mặt tế bào, đặc điểm này giúp các tế bào phân biệt chủng loại. Thí dụ, trong môi trường cấy các tế bào gan và thận riêng rẻ được trộn đều, sau một thời gian các tế bào gan nhận ra các tế bào cùng loại và tổ hợp lại với nhau; tương tự điều này cũng xảy ra ở những tế bào thận. Trong môi trường nuôi cấy tế bào, khi các tế bào va chạm nhau, tế bào ngừng chuyển động và sự tăng trưởng chậm và dừng lại, hiện tượng này gọi là sự ức chế do tiếp xúc (contact inhibition). Các sinh vật ký sinh cũng nhờ các carbohydrat trên glycocalyx để nhận ra tế bào chủ để xâm nhập vào. Cơ chế nhận dạng ở mức phân tử này cũng thường gặp như trường hợp các phản ứng dị ứng với các chất lạ cũng như trong sự dẫn truyền các xung thần kinh từ tế bào này sang tế bào khác.
![]()
6. Tiêm mao và chiên mao (cilia và flagella) |
Ở
một số tế bào động vật và tế bào
thực vật có một hay nhiều sợi tơ
giống như tóc cử động được
ở bề mặt ngoài của tế bào.
Nếu chỉ có một vài sợi và có chiều
dài tương đối dài hơn so với chiều dài
của tế bào thì được gọi là chiên mao,
nếu có rất nhiều và ngắn thì được
gọi là tiêm mao. Ở
tế bào chân hạch, cấu trúc cơ bản của
chúng giống nhau, và hai từ này dùng lẫn lộn qua
lại. Cả hai cơ
cấu này đều giúp cho tế bào chuyển
động hoặc làm chuyển động
chất lỏng quanh tế bào.
Thường gặp hai cơ cấu này ở
những sinh vật đơn bào và những sinh vật
đa bào có kích thước nhỏ và ở giao tử
đực của hầu hết thực vật và động
vật. Chúng cũng
hiện diện ở những tế bào lót ở
mặt trong của các ống, tuyến ở động
vật, các tiêm mao này giúp đẩy các chất di
chuyển trong lòng ống. Ở
khí quản người,
có thể có đến một tỉ tiêm mao trên 1cm2.
Tiêm mao và chiên mao của tế bào chân hạch là phần kéo dài ra của màng tế bào, với mười một nhóm vi ống nằm trong chất căn bản là tế bào chất; trong đó có chín nhóm với từng đôi vi ống xếp ở chu vi và hai vi ống đơn ở trung tâm (Hình 21). Mỗi chiên mao hay tiêm mao có một thể gốc ở đáy.
Tiêm
mao và chiên mao cử động khi vi ống này trượt
qua một vi ống khác. Các
vi ống ở ngoại biên được nối
với nhau bằng các tay protein, các protein này hoạt
động như các cầu bắt ngang giữa các
đôi vi ống. Khi các
tay protein uốn cong lại sẽ đẩy đôi vi
ống này qua một vị trí khác, sau đó các tay
rời ra và bắt đầu bám vào vị trí mới
trên đôi vi ống, và quá trình này được
tiếp tục gây ra sự cử động của tiêm
mao và chiên mao. Sự
cử động này cần năng lượng do ATP
cung cấp.
 |
Hình 21. Cấu tạo của tiêm mao và sự cử động của chúng
III.
CẤU TRÚC CỦA TẾ BÀO SƠ HẠCH
Hầu
hết những bào quan hiện diện trong tế bào chân
hạch thì lại không có trong tế bào sơ hạch.
Tế bào không có màng nhân và cũng không có các cấu
trúc có màng khác như mạng nội chất, hệ
Golgi, tiêu thể, peroxisom và ty thể (các chức năng
của ty thể được thực hiện ở
mặt trong màng của tế bào vi khuẩn).
Ở các vi khuẩn quang tổng hợp, có
những phiến hay túi có chứa diệp lục
tố mà không phải là các lạp có màng bao riêng
biệt.
Từ
lâu người ta nghĩ rằng tế bào sơ
hạch không có nhiễm sắc thể.
Hiện nay, với kính hiển vi điện
tử người ta biết rằng mỗi tế bào sơ
hạch có một phân tử ADN to,
mặc dù nó không liên kết chặt chẻ
với protein như ADN ở tế bào chân hạch, nó
vẫn được xem là nhiễm sắc thể.
Tế bào vi khuẩn thường có các ADN
nhỏ, độc lập được gọi là
plasmid. Nhiễm sắc
thể của tế bào sơ hạch và plasmid có
kiến trúc vòng kín.
Như nhiễm sắc thể của tế bào chân hạch, nhiễm sắc thể của tế bào sơ hạch có mang các gen kiểm soát các đặc điểm di truyền của tế bào và các hoạt động thông thường của nó. Sự tổng hợp protein được thực hiện trên ribô thể, một bào quan quan trọng trong tế bào chất ở cả tế bào sơ hạch và chân hạch. Tuy nhiên, ribô thể của tế bào sơ hạch hơi nhỏ hơn của tế bào chân hạch.
Một
số tế bào vi khuẩn có tơ mà quen gọi là chiên
mao. Những chiên mao này
không có vi ống và cấu trúc cũng như sự
cử động hoàn toàn khác chiên mao của tế bào
chân hạch. Chiên mao
của vi khuẩn chỉ được cấu tạo
bởi một loại protein
flagellin xếp theo
đường xoắn. Sự
cử động của vi khuẩn là do sự quay tròn
của các sợi protein này và đẩy tế bào
đi tới; sự đổi hướng di chuyển
là nhờ sự đảo ngược chiều quay
một cách tạm thời của các sợi protein
của chiên mao.
Vách tế bào của hầu hết vi khuẩn
được cấu tạo bởi murein gồm
một đường đa liên kết với các nhánh
acid amin, chỉ được tìm thấy ở tế bào
vi khuẩn.
*
Thuyết nội cộng sinh
Hiện
nay, các nhà sinh vật học cho rằng ty thể và
lục lạp là hai bào quan chỉ có trong tế bào chân
hạch có nguồn gốc từ những sinh vật sơ
hạch sống nội sinh trong tế bào sinh vật
chủ. Lynn Margulis
ở Ðại học Boston đưa ra các bằng
chứng về thuyết nội cộng sinh (endosymbiotic
hypothesis) như sau:
-
Ty
thể và lục lạp đều có chứa ribô
thể và nhiễm sắc thể riêng.
Ty thể và lục lạp cũng có thể
tạo màng riêng cho nó.
-
Nhiễm
sắc thể của những bào quan này giống
của tế bào sơ hạch, cả hai đều có
dạng vòng, không liên kết với thể nhân protein,
và không có màng bao.
-
Các
tổ chức bên trong của gen của các bào quan
giống như ở tế bào sơ hạch và rất
khác với tế bào chân hạch.
-
Ribô
thể của ty thể và lục lạp giống ribô
thể của tế bào sơ hạch hơn là tế bào
chân hạch.
-
Nhiều
vi khuẩn quang tổng hợp ngày nay sống bên trong
tế bào chân hạch, cung cấp chất dinh dưỡng
cho tế bào, ngược lại tế bào cung cấp nơi
ở cho chúng. Tương tự, một số vi
khuẩn không quang tổng hợp sống bên trong tế
bào chân hạch, lấy chất dinh dưỡng và chia
xẻ năng lượng từ thức ăn mà tế
bào chủ không thể biến dưỡng được,
do đó hình thức cộng sinh này là bắt buộc.
-
Sự
phân chia tế bào chân hạch có sự tham gia của
thoi vi ống trong khi vi khuẩn, ty thể và lục
lạp phân chia bằng cách phân đôi.
Theo
thuyết này, người ta giả sử rằng cách
đây 1,5 tỉ năm một tế bào đã bắt
được một vi khuẩn bằng cách nội
nhập bào. Vi khuẩn
chống lại sự tiêu hóa, sống cộng sinh bên
trong tế bào chủ và phân chia độc lập
với tế bào chủ. Sau
đó, một số gen của vi khuẩn xâm nhập vào
trong nhân của tế bào chủ và cùng khống
chế tế bào chủ. Theo
kiểu giải thích này thì tất cả những sinh
vật hiện nay có chứa ty thể, kể cả
thực vật, động vật, nấm và nguyên sinh
động vật. Cũng
theo thuyết này, các vi khuẩn quang tổng hợp khác
cũng có cùng nguồn gốc, kết quả là lục
lạp xuất hiện ở thực vật và tảo.
![]()
IV.
CÁC ÐẠI PHÂN TỬ QUAN TRỌNG TRONG TẾ BÀO
a.
Carbohydrat
Carbohydrat
là những hợp chất hữu cơ dồi dào
nhất trong tự nhiên. Chúng
là nguồn năng lượng hoá học cho cơ
thể sống, công thức chung là (CH2O)n; đơn
vị căn bản là những đường
đơn. Những carbohydrat đơn giản hiện
diện với tỉ lệ hai nguyên tử H, một
nguyên tử O (giống như nước) nối
với một C. Một
vài carbohydrat như tinh bột và celluloz là những đại
phân tử và rất
phức tạp, gồm nhiều đơn vị liên
kết nhau.
*
Ðường đơn (monosaccharide):
những phân tử carbohydrat căn bản là
những đường đơn. Các đường
đơn chứa ít nhất 3C, nhưng chúng có thể
chứa 5, 6 hay nhiều hơn.
Các đường 3, 5C có vai trò đặc
biệt trong tế bào sẽ được đề
cập trong các chương sau, còn đường 6C
nguồn cung cấp năng lượng cho tế bào là
những phân tử đường đơn để
tạo ra các đường phức tạp khác.
Có rất nhiều loại đường 6C, trong
đó glucoz và fructoz là hai loại đường quan
trọng nhất. Glucoz
là sản phẩm đầu tiên của quá trình quang
hợp và trở thành nguồn nguyên tử C ở mô
thực vật và động vật.
Ðường đơn có thể được
xếp loại theo số nguyên
tử carbon. Trioz
(glyceraldehyd) 3 C, pentoz (riboz) 5C và hexoz (như glucoz, fructoz và
galactoz) 6C. Fructoz và
galactoz là đồng phân của glucoz.

Hình 22.
Các loại đường đơn
Ðường đơn có thể ở dạng dây thẳng, nhưng trong nước chúng luôn luôn ở dạng vòng theo một trong hai cách. Dạng alpha, vị trí của gốc hydroxyl gắn với C thứ 1 ở phía dưới của mặt phẳng của vòng. Dạng beta, nhóm hydroxyl nằm trong mặt phẳng của vòng (Hình 23). Sự khác biệt giữa hai dạng tuy ít nhưng rất có ý nghĩa trong sinh học khi chúng trùng hợp để tạo ra các đại phân tử. Thí dụ: trùng hợp của (-glucoz tạo ra tinh bột và trùng hợp của (-glucoz tạo ra celluloz.

a-glucoz
b-glucoz
Hình
23. Các dạng của
glucoz
* Ðường đôi (disaccharide): gồm hai phân tử đường đơn nối với nhau bằng nối glycosidic, C thứ nhất của một phân tử đường đơn phản ứng với một gốc hydroxyl của một phân tử đường khác và mất một phân tử nước (Hình 24). Các đường đôi thông thường như sucroz, maltoz và lactoz. Sucroz (Hình 24) gồm hai phân tử glucoz và fructoz, và là dạng glucoz vận chuyển trong cây từ nơi quang tổng hợp ở lá đến những nơi khác trong cây. Maltoz gồm hai phân tử glucoz có nhiều ở lúa mì, lúa mạch được dùng làm rượu bia. Lactoz là đường đôi gồm glucoz và galactoz và là dạng đường của động vật hữu nhũ tạo sữa cho con. Ngoài ra ở côn trùng đường được vận chuyển trong haemolymph (máu) là trehaloz, một đường đôi gồm hai phân tử glucoz.

*
Ðường đa (polysaccharide): gồm nhiều
đường đơn nối với nhau
thành dạng thẳng hay phân nhánh, chúng có thể
được tạo thành từ một loại
đường, hai loại đường hay nhiều
hơn nữa. Chúng thường
không có vị ngọt, là những hợp chất không
tan với trọng lượng phân tử cao, và
được giữ ở dạng dự trử hay có
chức năng cấu trúc.
Tinh bột là một loại đường đa
dự trử chính của thực vật bậc cao
được hợp thành do amyloz và amylopectin (Hình 25A và
B), amyloz gồm hàng trăm phân tử glucoz nối nhau thành
chuỗi dài không phân nhánh. Mỗi
nối glycosidic được thành lập giữa C
thứ nhất của phân tử glucoz này với C
thứ tư của phân tử khác, nối (-1,4 (Hình
25A). Phân tử amyloz có
xu hướng cuộn tròn lại trong nước do
đó không tan (Hình 25C). Amylopectin
là những đường đa phân nhánh, dây ngắn
khoảng 20 - 30 phân tử glucoz được nối
nhau bằng nối (-1,4, ở vị trí nhánh gắn vào
dây chính bằng nối (-1,6 (Hình 25B).
Các dây nhánh giúp chúng không tan và không đứt
đoạn. Tinh bột
khoai tây gồm 20% amyloz và 80% amylopectin.
Inulin, chất dự trử ở các củ của
cây họ Cúc (Asteraceae), gồm những đơn
vị fructofuranoz được nối với nhau
bằng những nối glycosidic.

Hình 25A.
Amyloz
Hình 25B. Amylopectin

Hình
25C. Dạng xoắn
trong nước của amyloz
Hình
25. Các thành phần
của tinh bột
Glycogen
là những đường đa dự trử chính
của động vật, chúng có cấu trúc
tương tự amylopectin, nhưng phân nhánh nhiều hơn,
với khoảng 16 - 24 đơn vị (-glucoz của nhánh
bên cách nhau khoảng 8 - 10 phân tử glucoz của dây chính.
Ðường đa phong phú nhất là celluloz
được tìm thấy ở thực vật. Gỗ
của thực vật gồm 50% là celluloz còn ở
sợi bông vải hầu như 100% là celluloz.
Celluloz là những đường đa dây
thẳng gồm các đơn vị glucoz nối nhau
bằng nối (-1,4 (Hình 26), không giống tinh bột và
glycogen có các nối (-1,4. Celluloz
chỉ được thủy phân khi có enzim cellulaz xúc
tác, enzim này chỉ có ở một số vi khuẩn,
nguyên sinh động vật và nấm, không có ở
động vật. Một
số động vật có thể tiêu hóa được
celluloz nhờ các sinh vật cộng sinh trong ruột
của chúng như Kangaroo có vi khuẩn và mối có
những nguyên sinh động vật sống trong
ống tiêu hóa của chúng.

Hình
26. Celluloz
Thực vật còn có chứa những đường
đa có cấu trúc khác như pectin (gồm arabinoz,
galactoz và acid galacturonic) và hemicelluloz.
Chitin là thành phần cấu tạo của bộ xương
ngoài của côn trùng, giáp xác và vách tế bào nấm.
Chitin là một dạng biến đổi của
celluloz, trong đó một gốc có chứa nitơ
được thêm vào mỗi đơn vị glucoz
(N-acetyl-D glucosamin ) (Hình 27).

Hình
27. Chitin
b.
Lipid
Lipid
có nhiều chức năng sinh học quan trọng, chúng
là thành phần cấu trúc màng, mỡ và dầu có vai
trò dự trử và có thể tự cô lập.
Chúng làm thành cái áo bao bọc và bảo vệ cho
sinh vật không thấm nước ở bề mặt
ngoài và đôi khi hoạt động như một
yếu tố liên lạc về mặt hóa học
cả bên trong tế bào và giữa các tế bào.
Lipid
không tan trong nước do được cấu tạo
bằng những nối C - H không phân cực, chúng ngăn
cản các phân tử nước xâm nhập vào và
kết quả là những phân tử lipid kết tụ
lại với nhau tạo ra những giọt không tan.
Tuy nhiên, chúng dễ dàng tan trong các dung môi hữu
cơ như chloroform. Lipid
cũng gồm C, H, O nhưng O rất ít, ngoài ra, chúng còn
có chứa những nguyên tố khác đặc biệt
là P và N. Các loại
lipid khác nhau là: mỡ, dầu, sáp, phospholipid, glycolipid và
steroid.
*
Lipid dự trử: dầu và mỡ
Cùng
một đơn vị trọng lượng, dầu và
mỡ chứa nhiều năng lượng hơn
carbohydrat do chứa nhiều các nối C - H, trung bình 1 gr
mỡ chứa năng lượng gấp 2 - 3 lần
nhiều hơn carbohydrat hay protein.
Tất cả mỡ đều có sườn chính
là glycerol, chứa 3 nguyên tử C, mỗi phân tử
nối với một chuỗi acid béo tạo ra một
triglycerid (Hình 28). Acid béo
là một chuỗi những hydrocarbon với chiều dài
thay đổi và tận cùng là nhóm carboxyl (-COOH), các
acid béo được nối với phân tử glycerol
ở nhóm carboxyl; các acid béo trong dầu và mỡ ăn
được chứa từ 4 - 24 nguyên tử C.
Ba loại acid béo chính là acid stearic (18 C), acid palmitic
(16 C) và acid oleic (18 C). Acid
béo có số lượng H tối đa được
gắn vào C (như acid stearic) được gọi là
bảo hòa vì không có nối đôi giữa hai nguyên
tử C như bơ, mỡ heo và mỡ các động
vật khác. Acid béo
với các nối đôi được gọi là chưa
bảo hòa như dầu olive, dầu bắp, thường
ở dạng lỏng ở nhiệt
độ thường.
Dầu có hơn một nối đôi được
gọi là nhiều lần chưa bảo hòa
(polyunsaturated). Margarin chưa
bảo hòa chứa 70% của dầu nhiều lần chưa
bảo hòa có xu hướng được thay thế bơ
trong bửa ăn hàng ngày của những người
ăn kiêng vì các mỡ bảo hòa gây các bệnh
về mạch máu.
 |
Hình
28. Cấu tạo
của lipid
*
Lipid bảo vệ: sáp (waxe)
Sáp
tương tự như mỡ chỉ khác là acid béo
được nối với một chuỗi alcol dài hơn
glycerol, sáp không hòa tan trong nước, không có nối
đôi trong chuỗi carbon và trơ
(inert) đối với các phản ứng hóa học,
chúng làm thành lớp bảo vệ bề mặt ngoài
của nhiều động vật bao gồm cả
bộ xương ngoài của côn trùng, lông vũ
của chim và lông mao của thú và cả lớp
biểu bì của lá và trái của thực vật.
*
Lipid màng: phospholipid và glycolipid
|
|

+
Phospholipid: tương tự dầu và mỡ chỉ khác
là chúng có hai phân tử acid béo gắn vào mỗi phân
tử glycerol và một nhóm phosphat gắn vào C thứ ba
(Hình 29). Vì gốc
phosphat mang điện tích âm nên đầu phosphat
của phospholipid ưa nước (hydrophilic) và tan trong nước,
đuôi acid béo kỵ nước (hydrophobic), không tan
trong nước. Sự
khác biệt về tính chất đối với nước
của hai đầu của phân tử (nên có tên là lưỡng
ái: amphipatic) giữ cho
Hình 29. Phospholipid
phospholipid dễ dàng tạo thành màng đôi trong
cấu trúc tế bào. Khi
phospholipid được đưa vào nước, đầu
kỵ nước bị đẩy ra khỏi nước
và tụ tập lại với nhau tạo ra các micel hay
màng hai lớp lipid.
+
Glycolipid: là lipid có đường
cũng là thành phần quan trọng trong màng tế bào,
giống phospholipid có hai acid béo gắn vào phân tử
glycerol, nhưng có một dây carbon ngắn từ 1- 15
đường đơn nối vào C thứ ba.
Glycolipid cũng lưỡng ái và tạo thành micel
khi gặp nước tương tự phospholipid.
* Steroid:
cấu
trúc phân tử của steroid hoàn toàn khác các lipid
khác. Steroid có cấu trúc
gồm 4 vòng carbon, dẫn xuất từ cholesterol có
thể thêm vào một số đuôi hydrocarbon khác nhau (Hình
30).

Cholesterol
Hình 30. Steroid
Testosterol
![]()
Steroid
là những phân tử quan trọng trong động
vật có xương sống, chúng được tìm
thấy ở tim, mạch máu và gan.
Một số hormon và vitamin cũng là steroid.
Một trong steroid nhiều nhất là cholesterol, là
thành phần cấu tạo màng của tất cả
tế bào động vật và là điểm khởi
đầu để tổng hợp ra các hormon.
Cholesterol, không tan trong nước, có thể kết
tủa bởi mật trong túi mật, tạo ra sỏi
mật. Chúng tích
tụ trong các động mạch làm tăng huyết áp
và nguy cơ suy tim và nhồi máu cơ tim ở người
và những động vật khác.
Mức độ cholesterol trong máu không nhất
thiết là do chế độ ăn uống của con
người mà tùy thuộc nhiều hơn vào tỉ
lệ tổng hợp cholesterol của cơ thể.
c.
Protein
Protein
có cấu trúc phân tử rất thay đổi và có
rất nhiều chức năng: là enzim cho các quá trình
biến dưỡng, protein cấu trúc, cử động,
tồn trử, dinh dưỡng và vận chuyển, kháng
thể và toxin để bảo vệ và tấn công, và
hormon để điều hòa các chức năng
của cơ thể. Protein
chứa C, H, O, N và thường có S.
Ðơn vị cấu tạo protein là acid amin.
Chỉ có 20 loại acid amin khác nhau được
tìm thấy trong protein. Tuy
nhiên, số tổ hợp của acid amin sẽ tạo
ra rất nhiều loại protein.
* Acid amin:
tất
cả acid amin có cấu trúc tương tự nhau,
mỗi acid
amin có chứa một gốc amin ( - NH2, trừ prolin có
chứa một gốc imino - NH -), và một gốc
carboxyl (COOH), một nguyên tử H và một chuỗi bên
là gốc R gắn vào C trung tâm.

Hình 31. Cấu
tạo của 20 loại acid amin
Các
acid amin chỉ khác nhau ở gốc R, có thể chỉ
là một nguyên tử H như ở glycin, hay phức
tạp như cấu trúc có hai vòng như ở tryptophan
(Hình 31). Ðặc điểm
của mỗi acid amin ảnh
hưởng lên hình dạng của phân tử protein là
do gốc R mà ra.
Có
ba acid amin có chức năng đặc biệt là:
methionin là acid amin đặc biệt được
tổng hợp đầu tiên trong quá trình tổng
hợp protein, prolin làm cho chuỗi acid amin xoắn
lại và cystein nối các chuỗi acid amin lại
với nhau. Một
số gốc R phân cực (ưa nước), một
số khác thì không. Sự
không phân cực của chuỗi bên làm cho protein không
tan trong nước. Ðầu
tiên các acid amin được viết tắt bằng ba
chữ cái, bây giờ thì chỉ một chữ cái
(alanin, ala, A) ( Hình 31).
* Polypeptid:
một
chuỗi acid amin được gọi là polypeptid.
Acid
amin trong chuỗi polypeptid được nối với
nhau bằng liên kết peptid, được tạo ra
khi gốc - COOH của một acid amin này gắn với
gốc - NH2 của acid amin khác và giải phóng một phân
tử nước. (Hình 32). Khi
các acid amin liên kết nhau để tạo ra polypeptid luôn
luôn có một gốc amin tự do ở một đầu
N và một gốc carboxyl ở C tận cùng của
đầu kia, một protein có thể gồm một hay
nhiều sợi polypeptid với hàng trăm acid amin theo
một trình tự đặc biệt.
Các sợi polypeptid có thể liên kết chéo
với cầu nối cộng hóa trị là các cầu
nối disulfid, được
tạo ra giữa hai nguyên tử S của hai acid amin
cystein. Cầu nối
disulfide có thể nối các cystein trong cùng một
chuỗi polypeptid hay giữa các chuỗi polypeptid khác
nhau, kết quả là tạo ra cấu trúc không gian ba
chiều của protein.
![]()
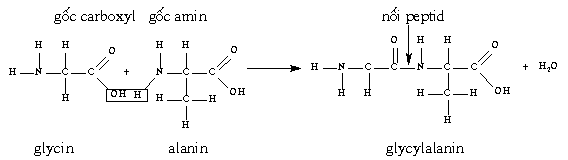
Hình
32. Sự thành lập
nối peptid giữa hai acid amin
*
Cấu trúc của protein
Protein
được chia làm hai loại chính: đơn
giản và phức hợp. Protein
đơn giản chỉ chứa acid amin.
Protein phức hợp (conjugated protein) gồm acid amin
và các thành phần hữu cơ hay vô cơ khác, thí
dụ: nucleo-, lipo-, phospho- và glycoprotein.
Những thành phần không phải là acid amin
của protein phức hợp được gọi là
gốc prosthetic.
Cấu trúc cấp một (sơ cấp) của
một protein là do trình tự thẳng của các acid
amin trong chuỗi polypeptid (Hình 33A).
Mỗi loại protein có cấu trúc cấp một
khác nhau.
Cấu trúc cấp hai của protein là do các cầu
nối hydro giữa các acid amin nằm kế nhau trong
cấu trúc cấp một (Hình 33B).
Thí dụ một cấu trúc cấp hai xoắn
phải (giống như một cầu thang xoắn.
Cấu trúc cấp ba là do các polypeptid có cấu trúc
cấp hai xếp chồng lên nhau, tạo ra một
khối hình cầu (Hình 33C).
Sự cuộn lại thường do sự tương
tác giữa các acid amin tương đối xa nhau trong.
Hình dạng cuối cùng cũng do ảnh hưởng
của môi trường,
đặc biệt là pH, do những vùng có điện tích
trên phân tử cũng ảnh hưởng lên hình
dạng của phân tử.

A
B
C
D
Hình
33. Bốn kiểu
cấu trúc của protein
|
|

Hình
dạng của phân tử quan trọng đặc
biệt ở những protein như hormon, là những thông
tin hóa học chỉ nhận protein đúng hình dạng
tương thích với nó như kiểu chìa khóa và
ổ khóa. Một số protein có kích thước
lớn như hemoglobin, chứa hơn một chuỗi
polypeptid, có cấu trúc khác hơn, cấu trúc cấp
bốn (Hình 33D). Hầu
hết những protein có kích thước lớn đặc
biệt là enzim có chứa hai sợi polypeptid hay hơn
nữa.
Hình dạng không gian ba chiều của protein
được gọi là dạng hay cấu hình của
protein. Tùy theo dạng
protein được chia ra làm hai loại chính: protein hình
sợi và protein hình
cầu. Trong protein hình
sợi, các sợi polypeptid xếp song song nhau theo
một trục đơn tạo ra các sợi hay bản
(collagen, keratin, elastin). Protein
hình sợi (Hình 34) không tan trong nước và thường
rắn chắc và là thành phần cấu tạo chính
của tóc, móng, sừng và vuốt.
Ở protein hình cầu, các sợi polypeptid cuộn
chặt thành hình tròn hay hình cầu.
Protein hình cầu tan trong nước và gồm
hầu hết các protein chức năng như enzim và các
hormon (Hình 33D).
Hình 34. Protein hình sợi
2. Enzim |
|
Các
phản ứng hóa học xảy ra trong tế bào
sống được tăng tốc nhờ những
chất xúc tác sinh học,
được gọi là enzim, và thường là các
protein. Một tế bào
động vật, có thể chứa đến 4.000
loại enzim khác nhau; mỗi loại xúc tác cho một
phản ứng hóa học khác nhau.
Một số enzim chung cho nhiều loại tế bào,
trong khi một số khác chỉ chuyên biệt cho
một loại tế bào để thực hiện
một chức năng đặc biệt.
Trong một phản ứng có enzim, chất phản
ứng được gọi là cơ chất (substrate).
Hầu hết enzim có tính chuyên biệt và chỉ
phản ứng một cách chọn lọc trên một
loại cơ chất nào đó mà thôi.
Là chất xúc tác, enzim giảm năng lượng
kích động cần thiết cho một phản
ứng xảy ra trong điều kiện nhiệt độ
và sinh lý bình thường (thí dụ 370C ở hầu
hết hữu nhũ). Enzim
có thể mang hai chất lại với nhau nhờ
tạo ra sự phối hợp tạm thời giữa các
cơ chất hay tập trung vào một cầu nối
hoa học đặc biệt trên cơ chất (Hình 35).
Tất cả enzim được dùng đi dùng
lại.
Enzim có khả năng xúc tác siêu tốc.
Thí dụ, phản ứng giữa CO2
+ H2O tạo ra H2 CO3 phản ứng xảy ra rất
chậm, có thể chỉ tạo ra 200 phân tử acid
carbonic/1 giờ khi không có enzim.
Trong tế bào sống, phản ứng được
xúc tác với enzim carbonic anhydraz ước chừng có
600.000 phân tử acid carbonic/1 giây.
Phản ứng nhanh gấp 10 triệu lần.
Trong cấu trúc không gian ba chiều, sợi
polypeptid của enzim cuộn lại tạo ra một hay
nhiều rảnh trên bề mặt, là những vùng
đặc biệt mà cơ chất có thể gắn vào.
Vùng này được gọi là hoạt điểm
(active site) (Hình 35). Hoạt
điểm của mỗi enzim có hình dạng đặc
biệt và chỉ có một loại phân tử cơ
chất có thể gắn vào.
Hoạt điểm
không những là nơi cơ chất gắn vào mà hướng
cơ chất theo một hướng nhất định.
Hoạt điểm có thể phân cực hay không,
ưa nước hay kỵ nước nên nó gắn
một cách chính xác vào phần nào của cơ
chất. Thí dụ
nếu ở phần phân tử cơ chất có điện
tích dương thì hoạt điểm của enzim
sẽ có điện tích âm.
Hoạt điểm của enzim chỉ gồm vài
acid amin của toàn thể enzim, những acid amin này
kề cận nhau trong một chuỗi polypeptid hay ở
những phần khác nhau của các chuỗi polypeptid nhưng
kề cận nhau do sự cuộn lại tạo ra
cấu trúc không gian ba chiều cấp ba và cấp
bốn của enzim. Gốc
R của acid amin được đưa ra ngoài tạo
ra hoạt điểm và cách sắp xếp của chúng
quyết định sự gắn cơ chất với
enzim.

Hình
35. Mô hình cơ chế
xúc tác của enzim
*
Mô hình hoạt động của enzim
+
Mô hình ổ và chìa khóa (Hình
36)
Theo
thuyết này, một cơ chất gắn vào hoạt
đểm như một chìa khóa gắn khít vào ổ,
cơ chất và enzim gắn vào nhau tạo ra một
phức hợp enzim-cơ chất.
Cơ chất bị
biến
đổi một cách chuyên biệt và kết quả là
sản phẩm được phóng thích.

Hình 36.
Mô hình ổ và chìa khóa
Hình 37. Mô hình
cảm ứng
+
Mô hình cảm ứng (the induced-fit model) (Hình 37)
Các
nghiên cứu cấu trúc của enzim cho rằng hoạt
điểm linh động hơn lỗ của ổ khóa.
Trong một số trường hợp, trong lúc cơ
chất gắn vào, cấu hình thay đổi do hình
dạng hoạt điểm của enzim bị cảm
ứng làm cho chúng khớp với nhau hơn.
Sự cảm ứng này làm thay đổi lực
giữa các nối đặc biệt của phân tử
phản ứng do đó phản ứng có xúc tác
được dễ dàng xảy ra.
*
Các yếu tố ảnh hưởng lên hoạt động
của enzim
+
pH có thể làm thay đổi cấu hình của enzim,
đặc biệt là sự tương tác giữa
những acid amin có điện tích âm (acidic) và điện
tích dương (basic). Khi
pH thay đổi, hình dạng của hoạt điểm
có thể thay đổi, do đó cơ chất không
thể gắn vào được.
Hầu hết enzim hoạt động tốt
nhất trong khoảng pH 6 - 8.
Tuy nhiên, pepsin giúp tiêu hóa protein
trong bao tử hoạt động trong khoảng pH
rất thấp (2). Pepsin
có một trình tự acid amin duy trì cầu nối hydro và
ion hóa ngay cả khi có sự hiện diện của acid
mạnh. pH cũng
ảnh hưởng trên phản ứng trong đó ion H+
là một thành phần của sự thay đổi hóa
học đượcû xúc tác.
+ Nhiệt độ thay đổi dễ làm đứt
các cầu nối hydro, các cầu nối này và sự tương
tác giữa các cực kỵ nước giúp giữ
cấu trúc cấp ba, cấp bốn của protein kể
cả enzim. Sinh vật
thường có những enzim hoạt động tối
ưu ở nhiệt độ mà chúng sống.
Ở hữu nhũ, nhiệt độ tối
ưu từ 35 - 400C. Dưới
nhiệt độ này, các nối để giữ hình
dạng của protein không đủ linh động
để thay đổi hình dạng cần thiết
tham gia phản ứng. Do
đó, enzim hoạt động không hữu hiệu
ở nhiệt độ dưới 350C. Trên
400C, các cầu nối hydro và các cầu nối yếu
khác không thể giữ protein ở
hình dạng chính xác của nó.
Một enzim bị biến tính một phần
bởi nhiệt có cấu trúc bị quéo lại một
chút và chuỗi polypeptid thay đổi cấu hình
(conformation) có thể trở lại hình dạng cũ
nếu được làm nguội, nhưng nếu
biến tính hoàn toàn như albumin của trứng
luộc chín thì không thể trở lại dạng ban
đầu được nữa.
*
Cofactor và coenzim
Nhiều
enzim cần phải có thêm các thành phần hóa học
khác để thực hiện chức năng, phần
thêm đó được gọi là cofactor.
Cofactor có thể la ìmột phân tử hữu cơ
hay các ion kim loại (sắt, mangan, kẻm...). Khi cofactor
là một phân tử hữu cơ không phải là
protein,û như một vitamin, thì được gọi là
coenzim. Tương
tự enzim, coenzim cũng được sử dụng
lại sau khi xúc tác phản ứng.
Dù cần với một lượng rất
nhỏ nhưng nếu thiếu coenzim thì rất tai
hại đối với đời sống sinh vật,
cho nên trong khẩu phần cần phải có vitamin.
Coenzim và cofactor được tìm thấy ở các
enzim trong các chuỗi dẫn truyền điện tử
của các quá trình quang hợp và hô hấp của
tế bào.
![]()